
Abstract
Objective To compare gut microbiome diversity and composition in SARS-CoV-2 polymerase chain reaction (PCR)-confirmed positive patients whose symptoms ranged from asymptomatic to severe versus PCR-negative exposed controls.
Design Using a cross-sectional study design, we used shotgun next-generation sequencing (NGS) to evaluate microbiome composition and diversity in both patients with SARS-CoV-2 PCR-confirmed infections presenting to Ventura Clinical Trials for care from March 2020 through January 2021 and SARS-CoV-2 PCR-negative exposed controls. Patients were classified as being asymptomatic or having mild, moderate, or severe symptoms based on NIH criteria.1 Exposed controls were individuals with prolonged or repeated close contact with patients with SARS-CoV-2 infection or their samples, e.g. household members of patients or frontline healthcare workers. Microbiome diversity and composition were compared between patients and exposed controls and across patient subgroups at all taxonomic levels.
Results There were 52 patients and 20 controls. Compared with controls, patients had significantly less bacterial diversity, a lower abundance of Bifidobacterium and Faecalibacterium as well as some other bacteria, and a higher abundance of Bacteroides at the genus level. Additionally, there was an inverse association between disease severity and both bacterial diversity and Bifidobacterium and Faecalibacterium abundance.
Conclusion We hypothesize that low bacterial diversity and depletion of Bifidobacterium and Faecalibacterium genera either before or after infection led to reduced pro-immune function, thereby allowing SARS-CoV-2 infection to become symptomatic. This particular dysbiosis pattern may be a susceptibility marker for severe symptoms from SARS-CoV-2 infection and may be amenable to pre-, intra-, or post infection intervention.
1. Introduction
Of all the bacteria colonizing the gastrointestinal tract, the Bifidobacterium genus within the Actinobacteria phylum is one of the most recognized and studied. They colonize the human gastrointestinal tract soon after birth, their relative abundance is inversely associated with both age and weight2 and species within this genus represent the active ingredient in many probiotics. Theorized benefits of these Gram positive bacteria include enhanced ATP production, immune modulation,3 mucosal barrier integrity, restriction of bacterial adherence to and invasion of the intestinal epithelium, and modulation of central nervous system activity.4,5
In vitro studies suggest that Bifidobacterium have immune-modulatory properties6,7-10 and may serve as a marker of high overall immunue competence.11 Many bifidobacteria have demonstrable anti-inflammatory properties: B. animalis, longum, and bifidum decrease the function of the “master switch”3 pro-inflammatory tumor necrosis factor-(TNF-) α, increase the anti-inflammatory cytokine interleukin-(IL-10), and promote TH1 while inhibiting TH2 immune response.11 In a mouse model of inflammatory bowel disease, B. bifidum and animalis reduced pro-inflammatory cytokines and restored intestinal barrier integrity.11 Relevant to diseases affecting the lungs, such as SARS-CoV-2 infection, there is immunologic coordination between the gut and lungs – the gut-lung axis.12-14
In the gut microbiota of SARS-CoV-2 infected patients, researchers have observed that increased abundance of the genera Streptococcus, Rothia, Veilonella, and Actinomyces was associated with inflammation, 15 increased abundance of Collinsella aerofaciens, Collinsella tanakaei, Streptococcus infantis, and Morganella morganii were associated with fecal samples with high SARS-CoV-2 infectivity,16 and increased Lachnospriaceae and Enterobacterioaceae abundance was associated with both increased need for artificial ventilation and mortality.16 Species mentioned as being potentially protective include Parabacteroides merdae, Bacteroides stercoris, Alistipes onderdonkii, Lachnospiracea bacterium16 and Faecalibacterium prausnitzii.16,17 At least two reports have described a decreased abundance of Bifidobacterium bifidum associated with SARS-CoV-2 infection and severity.17,18
The scientific community and lay public are increasingly interested in the therapeutic potential of probiotics. Bifidobacteria spp. have shown potential to address a number of clinically important entities ranging from inflammatory bowel disease (IBD)19 to infection caused by Clostridioidies difficile.19-22 Studies have shown that treatment with specific strains of Bifidobacteria helps restore colonic integrity in vivo23 and reduce both Clostridioidies difficile counts and toxin levels in vitro.21 There are also in vivo data suggesting that Bifidobacterium longum administered intranasally prior to exposure to influenza reduces mortality in mice.7
A number of studies have suggested that a healthy gut microbiome might be associated with decreased SARS-CoV-2 related mortality24 and that probiotics should be considered for prophylaxis25 or treatment of SARS-CoV-2 infections25 or as adjuvants to either therapy or vaccination.26 Others have suggested that probiotics might reduce the incidence of secondary infections resulting from bacterial translocation.27
As of May 2021, nearly 8000 studies had been registered worldwide to find therapies or vaccines for SARS-CoV-2 infection. Besides our current study, only two of these were focused on the effect of SARS-CoV-2 on the microbiome (clinicaltrials.gov: NCT04443075 and NCT04486482). In this study, we evaluate the relationships between got microbiome diversity and composition compared to clinical outcome in cross-sectional groups of SARS-CoV-2 PCR-confirmed positive patients (ranging from asymptomatic to severely symptomatic) versus SARS-CoV-2 PCR-confirmed negative controls. This study is unique in that it compares SARS-CoV-2 positive patients versus SARS-CoV-2 exposed (i.e., either non-quarantined household members or frontline healthcare workers) persons who remained PCR-negative and asymptomatic (“controls”). The controls had similar viral exposures, but appeared to be protected against infection. Our comparison of the microbiome in patient vs. control subjects suggests that at least some protection may reside in the microbiome.
2. Methods
2.1. Study Design and Patients
Patients diagnosed with SARS-CoV-2 infection who sought remote care from Ventura Clinical Trials in 2020 and 2021 were recruited to the study following expression of interest or physician referral. Patients eligible for enrolment were aged ≥18 years of age with RT-PCR-confirmed SARS-CoV-2 infection within one week of screening. Controls eligible for enrollment were PCR-negative for SARS-CoV-2, remained antibody negative for 3 to 6 months, and asymptomatic for 6 to 12 months. SARS-CoV-2 negative controls had to meet the following additional criteria: they either shared a household with at least one symptomatic SARS-CoV-2-positive family member (e.g., sibling, parent, or child) or were a healthcare worker who had been repeatedly exposed to symptomatic SARS-CoV-2-positive patients or numerous SARS-CoV-2 samples. Additionally, all exposed controls were ones that, despite exposure to SARS-CoV-2, chose not to quarantine or take prophylaxis for SARS-CoV-2 infection and none had yet been vaccinated. Of importance, patients with, or family members or staff exposed to SARS-CoV-2 did not wear PPE in the presence of their family or in doctor’s offices. Healthcare workers and staff did not wear full PPE (i.e., did not wear masks) because of their scarcity during this global pandemic. Patients undergoing treatment with total parenteral nutrition, or those with a history of significant gastrointestinal surgery (e.g. bariatric surgery, total colectomy with ileorectal anastomosis, proctocolectomy, postoperative stoma, ostomy, or ileoanal pouch) were excluded.
The study was conducted in accordance with ethical principles of the Declaration of Helsinki, the International Council for Harmonisation (ICH) Harmonised Tripartite Guideline for Good Clinical Practice (GCP), and was approved by the “Ethical and Independent Review Services” (https://www.eandireview.com/) Independent Review Board (IRB). All patients provided written informed consent to participate in the study and the study is registered as clinicaltrials.gov NCT04031469.
2.2. Assessments
A self-administered questionnaire solicited information on symptom severity, past medical history, current medication and probiotic use, and exposure to recreational drugs or animals. Patients with SARS-CoV-2 infection were further classified as being either asymptomatic carriers or having mild, moderate, or severe symptoms as per National Institute of Health, Clinical Spectrum of SARS-CoV-2 Infection criteria.1,28 PCR-confirmed SARS-CoV-2 positive household members of SARS-CoV-2 infected patients who lacked symptoms were categorized as asymptomatic carriers. Patients and controls were classified as underweight, normal weight, overweight, obese, or severely obese based on body mass index criteria of the Center for Disease Control and Prevention.29
2.3. Stool Sample Collection and Processing
Patients and controls within the same household collected stool samples within a week of the index case being positive. They were instructed to collect 1 mL of fresh stool and place it directly in a Zymo Research DNA/RNA Shield fecal collection tube. Following fecal collection, each individual sample DNA was extracted and purified with the Qiagen PowerFecal Pro DNA extraction kit. The isolated DNA was then quantitated utilizing the Quantus Fluorometer with the QuantFluor ONE dsDNA kit. After DNA quantification, the DNA was normalized, and libraries were prepared using shotgun methodology with Illumina’s Nextera Flex kit. Per our lab shotgun metagenomic processes, samples underwent tagmentation, amplification, indexing, and purification. After completing the NextSeq run, these raw data were streamed in real-time to Illumina’s BaseSpace cloud for conversion to FASTQ files. The FASTQ data were compared to evaluate microbiome diversity in patients with PCR-confirmed SARS-CoV-2 infections and SARS-CoV-2 PCR-negative exposed controls. We compared microbiome differences between the SARS-CoV-2 patients and SARS-CoV-2-negative exposed controls at all taxonomic levels.
2.4. Data Analysis
We assessed differences in relative abundance across taxa between the gut microbiome of SARS-CoV-2 infected patients and exposed controls and calculated Shannon and Simpson index with One Codex’s bioinformatics analysis pipeline using Jupyter notebook in Python. To evaluate the statistical significance of any variability noted between patients and exposed controls, t-test, ANOVA, and chi-square tests statistics were conducted using R version 3.6.1 (2019-07-05), GraphPad vs. 8, and SigmaPlot 12.0. P-values <0.05 were considered significant.
3. Results
3.1. Patient Characteristics
Demographic and clinical characteristics of patients (n = 52) and exposed controls (n = 20) are presented in Table 1. Half (50%) of patients and 35% of exposed controls were male. The median (SEM) age of patients was 53 ± 2.5 years and of exposed controls was 48 ± 3.6 years. A total of 88.5% of patients were non-Hispanic White; 9.6%, Hispanic; 1.9%, Native American and 85.0% of exposed controls were non-Hispanic White; 10.0%, Hispanic; 5.0%, Black. Of patients, 57.7% had severe disease, 23.1% had moderate disease and 11.5% had mild disease; the remaining 7.7% were asymptomatic. 78.9% of patients and 60.0% of exposed controls had underlying comorbidities considered risk factors for increased morbidity and mortality by the CDC.2 The mean ± SEM BMI of the 52 patients for whom data were available was 27.9 ± 0.94 compared with 25.1 ± 0.96 for the 20 exposed controls. There was no statistical difference (p>0.05) in gender, age, racial demographics, or presence of underlying comorbidity.
Demographic and baseline clinical characteristics of SARS-CoV-2-positive patients and SARS-CoV-2-negative exposed controls
Ranges: 1-5, 6-10, 11-15, 16-20, 21-25
Of the exposed controls, 16 were family members of various SARS-CoV-2 positive patients, 2 were healthcare workers with extensive, non-protected, exposure to subjects, and 2 were laboratory personnel exposed to thousands of SARS-CoV-2 samples (healthcare workers and laboratory personnel did not wear full PPE (i.e., did not wear a face mask) due its scarcity; see methods). During the timeframe of the study, none of the patients or controls were on SARS-CoV-2 prophylaxis or treatment, and none had yet been vaccinated.
3.2. Gut Microbiome Diversity and Composition
Alpha diversity in patients categorized as severely affected based on NIH criteria and SARS-CoV-2 negative controls are shown in Figure 1, with a significant decrease in Shannon Index (P=0.026) and Simpson Index (P=0.043) in relation to SARS-CoV-2 positivity, suggesting that the gut microbiome of SARS-CoV-2 infected patients was less diverse than that of exposed controls.
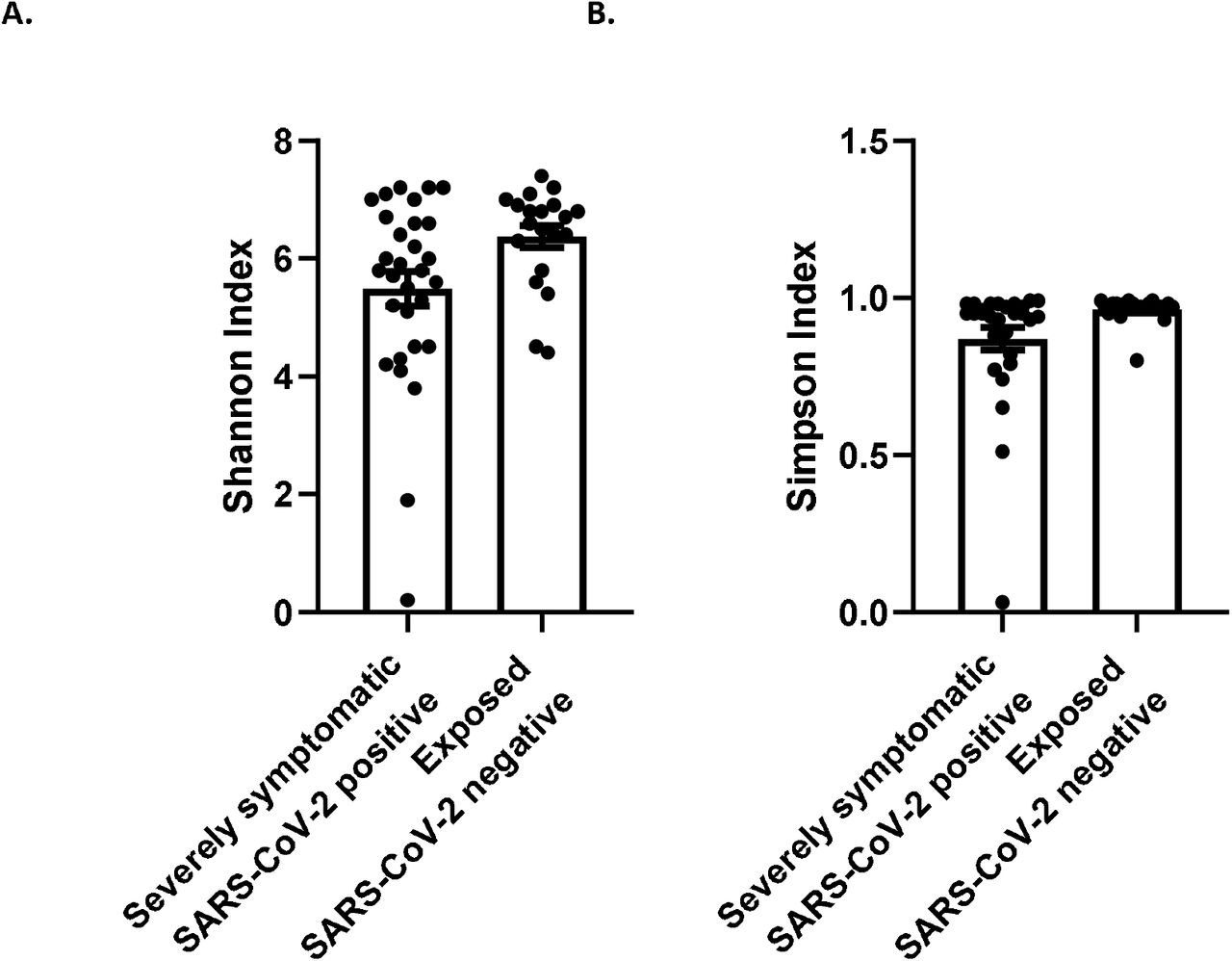
Alpha diversity of SARS-CoV-2 positive patients with severe symptoms (n=30) vs. SARS-CoV-2 negative exposed controls (n=20). A. Shannon index (P=0.026) B. Simpson index (P=0.043)
Microbiome analysis revealed significant differences between the SARS-CoV-2 patients and controls. Table 2 lists the bacteria that differed in abundance in relation toSARS-CoV-2 positivity (center column) and SARS-CoV-2 infection severity (right column) for all patients and controls. Patients with PCR-confirmed SARS-CoV-2 infection showed a significantly decreased abundance of Bifidobacterium, Clostridium, Faecalibacterium, Faecalibacterium prausnitzii, Ruminococcus, and Subdolingranulum and significantly increased abundance of Bacteroides (Figure 2, Table 2 middle column). Table 2 right column shows that bacterial abundance was associated with symptom severity. A one-way ANOVA analysis revealed a significant association between increased disease severity and decreased abundance of Bifidobacterium, Faecalibacterium, Faecalibacterium prausnitzii, Ruminococcus, and Subdolingranulum and increased abundance of Bacteroides and Dorea. Figure 2 summarizes Table 2, with green boxes depicting significant associations between changes in bacterial abundance and either SARS-CoV-2 positivity or SARS-CoV-2 infection severity.
Associations of and SARS-CoV-2 positivity (center column) and SARS-CoV-2 infection severity (right column) with bacterial abundance for all bacteria (left column) tested. Values indicate p-values determined by either one-way ANOVA (center column, analyzing all SARS-CoV-2 positive severities) or unpaired t-test (right column). Values of p<0.05 (bold) indicate an association. The direction of the arrows indicates whether the bacteria are decreased or increased with SARS-CoV-2 positivity or SARS-CoV-2 infection severity.
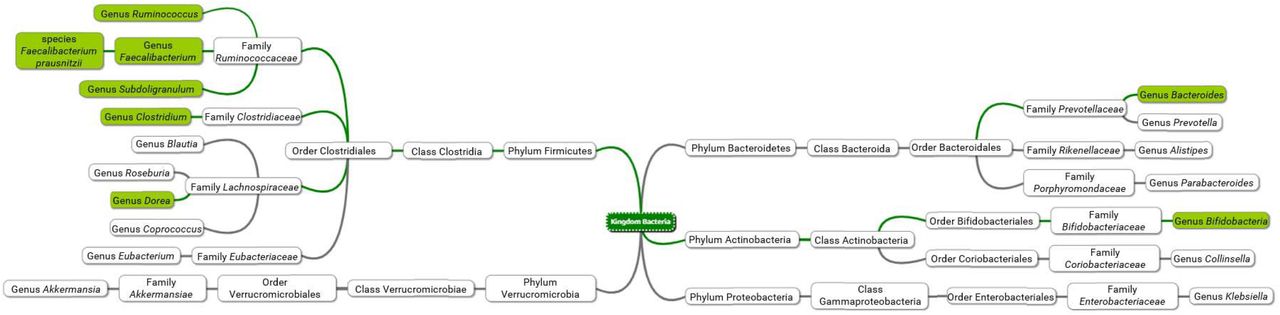
Diagram of taxa, comparing the gut microbiome of SARS-CoV-2 positive patients and SARS-CoV-2 negative exposed controls. Green background indicates a significant association between abundance for either SARS-CoV-2 positivity or SARS-CoV-2 infection severity.
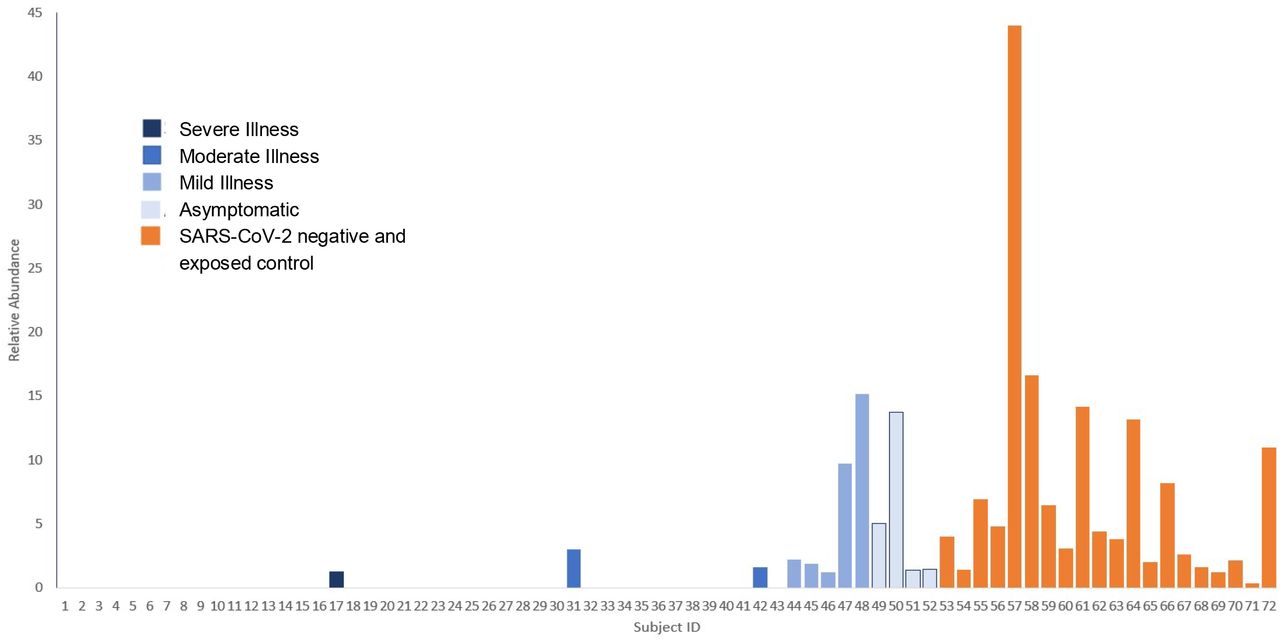
Figure 3 shows the relative abundance of Bifidobacterium for each subject, grouped by SARS-CoV-2 infection severity. Analyzed via one-way ANOVA with Dunnett’s post-hoc test (comparing all SARS-CoV-2 infection severities including asymptomatic to controls), there was a significant (P=0.0002) association between severity and Bifidobacterium relative abundance, particularly for severe (P<0.0001) and moderate (P=0.0031) severity infections.

Relative abundance of Bifidobacteria in SARS-CoV-2 positive patients compared to SARS-CoV-2 negative exposed controls, grouped by disease severity.
Figure 4 shows the top 18 most common families and genera for patients stratified by disease severity and exposed controls. This diagram presents relatively raw data of the abundance of various families and genera. In column B, for SARS-CoV-2 positive patients’ (top) microbiome, the reduced diversity compared to controls (bottom) can be subjectively observed.
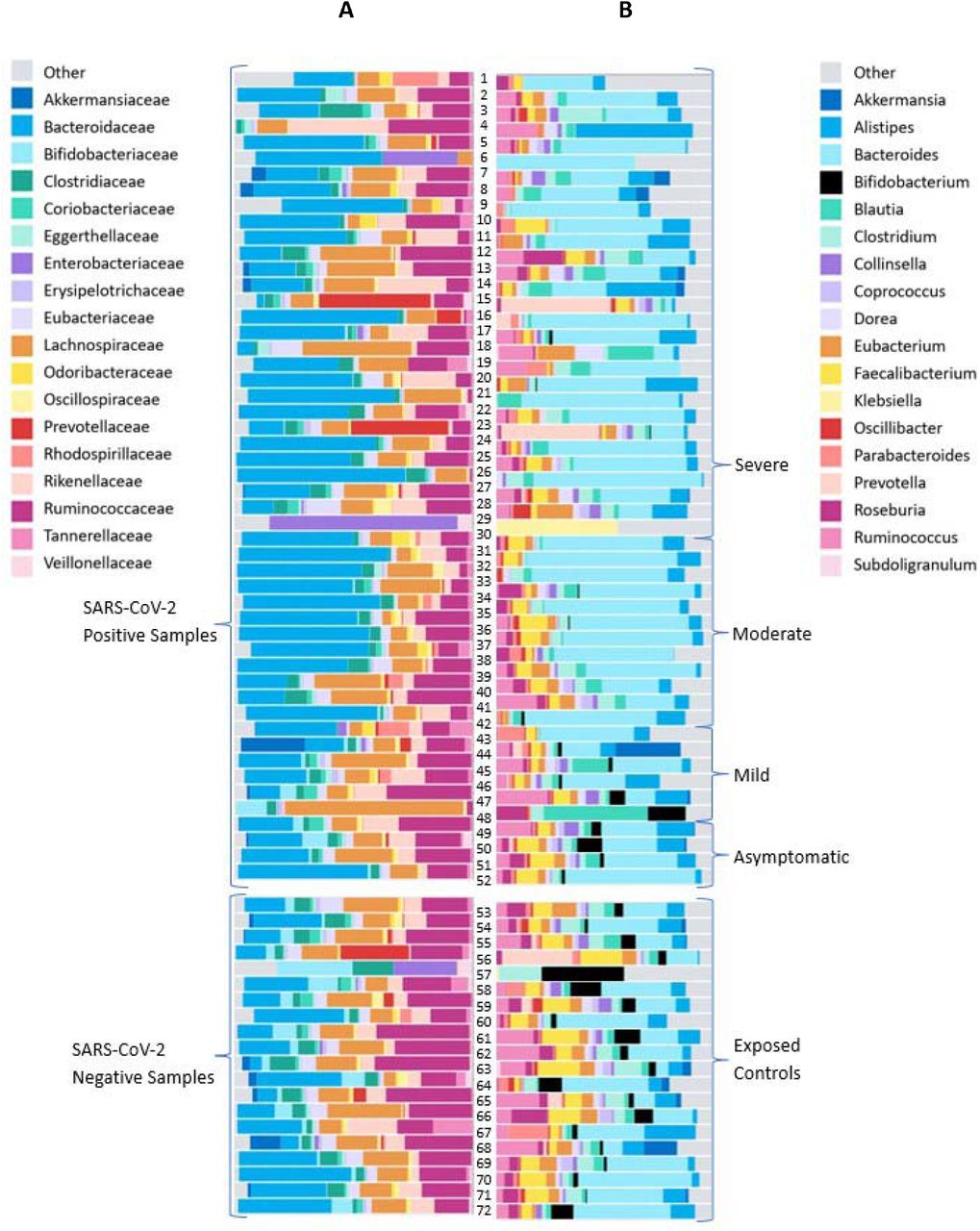
Relative abundance of the top 18 most common A. families and B. genera. The top group represents the SARS-CoV-2 positive samples, stratified by severity. The bottom group represents the exposed control samples.
4. Discussion
4.1. Immune function and health could be enhanced by bacterial abundance
Interactions between the host and gut microbiota are complex, numerous, and bidirectional. Gut microbiota regulate the development and function of the innate and adaptive immune systems,30 potentially allowing them to protect against both infections and infection severity. The primary findings of our study are that SARS-CoV-2 positivity and infection severity are associated with decreased levels of bacterial diversity and decreased levels of the protective Bifidobacterium and Faecalibacterium genera. Uniquely, our study compared SARS-CoV-2 exposed SARS-CoV-2 negative persons (i.e., controls) with symptomatic and asymptomatic SARS-CoV-2 positive patients. Thus, we controlled for SARS-CoV-2 exposure..
Bacterial diversity is known to inversely relate to presence of various common disorders.31 Our study had similar findings: higher diversity, as exemplified by the Shannon and Simpson indices, correlated (p<0.05) with reduced SARS-CoV-2 infection severity.
The genus Bifidobacterium has important immune functions,11 is a major component of the microbiome, and is frequently used in probiotics.32 Bifidobacteria are Gram positive non-motile anaerobic rod-shaped bacteria. They protect against intestinal epithelial cell damage and this protection is independent of their effects on tumor necrosis factor alpha (TNF-α) production. The exopolysaccharide coat which is a feature of some Bifidobacterium has been shown to play a significant role in this protective effect.33 Bifidobacteria also reduce cell damage by inhibiting TNF-α and macrophages, and they increase Treg responses.34 These immune functions of Bifidobacterium could be critical in relation to its SARS-CoV-2 infection-prevention effects.
Evidence has accumulated to support a beneficial effect from supplementation with Bifidobacterium in a number of disease states.39 The number of commensal bifidobacteria have been shown to decrease with age,39 a major SARS-CoV-2 infection risk factor. In our study, a significant relationship between disease severity and relative abundance of Bifidobacterium was observed; patients with a more severe course of viral infection decreased abundance of Bifidobacterium. However, it should be noted that there are no definitive studies concerning what constitutes a normal baseline abundance of Bifidobacterium in a “healthy” individual.
The abundance of Faecalibacterium genus and Faecalibacterium prausnitzii species were also inversely related to SARS-CoV-2 positivity and SARS-CoV-2 infection severity in this analysis. Age and diabetes are risk factors for SARS-CoV-2 infection, and Faecalibacterium prausnitzii levels decline markedly in elder and diabetic populations.35 In fact, Faecalibacterium levels have been considered an indirect “indicator” of overall human health.36 The “Western” diet (consumption of more meat, animal fat, sugar, processed foods, and low fiber) reduces levels of F. prausnitzii, while high-fiber (e.g. Mediterranean diet of vegetables and fruits) and low meat intake enhances the abundance of F. prausnitzii.37 Preliminary studies showed that reduced ingestion of a Mediterranean diet within the same country is associated with increased SARS-CoV-2 related death rates.38 In short, we show that F. prausnitzii levels negatively correlated to SARS-CoV-2 infection severity and prior studies show that reduced F. prausnitzii is associated with SARS-CoV-2 infection vulnerabilities such as age, diabetes, obesity, and possibly diet.
In addition to the negative correlations between relative abundance of Bifidobacterium and Faecalibacterium (especially F. prausnitzii) with both infection positivity and disease severity, we observed changes in the abundance of other characterized bacterial genera. SARS-CoV-2 positivity was associated with decreased abundance of Clostridium, Ruminococcus, and Subdolingranulum and increased abundance of Bacteroides. SARS-CoV-2 infection severity was associated with increased abundance of Bacteroides and Dorea. The implications of these changes remain unclear.
4.2. Innate immunity could be enhanced by increased bacterial level
The pathological impact of SARS-CoV-2 infection includes both direct effects from viral invasion and complex immunological responses including, in its most severe form, the ‘cytokine storm.’ The cytokine storm is the result of a sudden increase in circulating levels of pro-inflammatory cytokines produced by activated macrophages, mast cells, endothelial cells, and epithelial cells during innate immune responses, which appear to be modulated by the abundance of Bifidobacteria and Faecalibacteria and bacterial diversity.5,30,36 Steroid treatment has situational success in SARS-CoV-2 infection, which is based on suppressing this over activation of the innate immune system, as reviewed by Tang et al.40
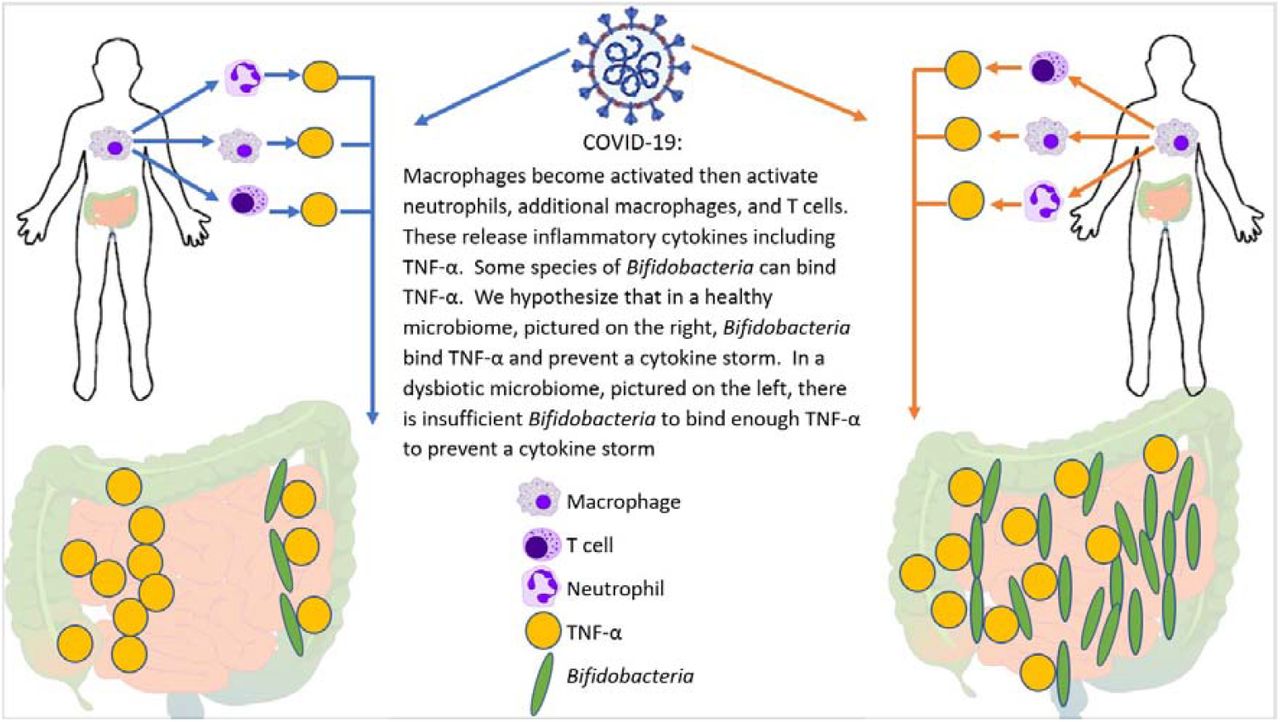
Zhao et al. reported that elevated serum levels of pro-inflammatory cytokines such as IL-16 and IL-17 predict poor prognoses in patients with SARS-CoV-2 infection.41 Also, Tao et al. showed that changes in gut microbiota composition might contribute to SARS-CoV-2-induced production of inflammatory cytokines in the intestine, which may contribute to the onset of a cytokine storm.28 Both authors report significantly reduced gut microbiota diversity, as well as increased opportunistic pathogens in SARS-CoV-2 patients. Interestingly, the bloom of opportunistic pathogens positively correlated with the number of Th17 cells. In another article, Bozkurt et al. reported that IL-6 and IL-17 promote viral persistence by immune interactions through cellular autophagy via the inositol-requiring enzyme 1 pathway.26 They also cited that some species of Bifidobacterium are likely to prevent the replication of coronaviruses by reducing endoplasmic reticulum stress induced through the inositol requiring enzyme 1 pathway. Alterations in gut microbiota and reduced Bifidobacterium levels have been previously reported in the gastrointestinal tract of patients with IBD.29 IL-17 also plays an important role in IBD. Furthermore, the direct endoscopic delivery of Bifidobacterium has been shown to be effective in promoting symptom resolution and mucosal healing―an effect likely to be associated with the anti-Th17 effect of Bifidobacterium. The findings reported in the current study support the role of commensal Bifidobacterium in the severity of viral infection, suggesting the importance of microbiota in viral pathogenesis and their potential therapeutic implications. Figure 5 demonstrates how bifidobacteria might quell a heightened immune response by dampening the effect of the master switch TNF-α.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Potential mechanism for cytokine storm and immune hyper-response in SARS-CoV-2 positive patients. In individuals infected with SARS-CoV-2, the macrophages become activated; these in turn activate T-cells, additional macrophages, and neutrophils―all of which release cytokines, including the “master switch” TNF-α. Bifidobacteria, when present in sufficient numbers, can bind to TNF-α and prevent the subsequent cytokine storm. Therefore, patients with a bifidobacterial dysbiosis characterized by low levels of Bifidobacteria lack this line of defense which may lead to a cytokine storm.
5. Conclusions
Given our cross-sectional study design, it is not possible to determine whether the differences in Bifidobacterium levels observed between patients and exposed controls preceded or followed infection. If the differences preceded SARS-CoV-2 infection and are a marker of susceptibility, then boosting Bifidobacterium levels might be expected to decrease the risk or severity of SARS-CoV-2 infection. If these changes followed SARS-CoV-2 infection, then adjunctive refloralization of the gut microbiome and boosting of Bifidobacterium through supplementation or fecal transplantation could speed recovery and reduce organ damage. Refloralization may be particularly helpful for SARS-CoV-2 infection “long haulers,” patients hospitalized with ‘severe’ disease (including children with SARS-CoV-2-related multisystem inflammatory syndrome) and those with persistent symptoms. Future studies of individuals with baseline pre-pandemic microbiome data would be highly valuable. However, acquiring baseline pre-infection microbiome data is not at present inexpensive. Developing outbreaks within tightly closed communities such as nursing homes might be a good setting in which to assess susceptibility: fecal samples could be collected during the outbreak and run post hoc on “cases” and “controls.”
SARS-CoV-2 infection presentation variability appears to show some correlation with colon microbiome bacterial composition and overall diversity. The same changes we observe due to SARS-CoV-2 infection, namely reduced Bifidobacterium and/or Faecalibacterium abundance, are likely associated with SARS-CoV-2 infection risk factors including old age, obesity, and diabetes.24,35,37,42 Thus, colon microbiome diversity and relative abundance of Faecalibacterium and Bifidobacterium may be markers that could be used to predict SARS-CoV-2 infection severity. Moreover, this suggests that evaluating an individual’s gut microbiome could provide important information for SARS-CoV-2 treatment or prevention. In summary, in this study, we present findings from SARS-CoV-2-exposed subjects that include both those who remained healthy as well as those who became ill at various levels of severity. Reduced bacterial diversity and reduced levels of various bacteria are significantly related to both SARS-CoV-2 positivity and SARS-CoV-2 infection severity. These findings suggest that Bifidobacterium and other probiotic supplementation or refloralization via fecal transplantation might provide a therapeutic benefit, particularly for patients hospitalized with severe disease. The data also suggest that individual colon microbiome evaluation may predict vulnerability to the development of severe SARS-CoV-2 infection. Lastly, our data suggest a new imperative: going forward, colon microbiome patterns should be controlled for in studies intended to identify preventive (e.g., vaccines), prophylactic, or therapeutic measures for SARS-CoV-2 infections.
Data Availability
Data available upon reasonable request from corresponding author, Dr. Sabine Hazan.
DECLARATIONS
Funding
No funding was received for this study. Several authors were compensated by ProgenaBiome.
Conflicts of Interest/Competing Interests
SH and TJB have corporate affiliation to ProgenaBiome and Topelia therapeutics. AJP and BDB have corporate affiliations to Progenabiome. SD has corporate affiliation to McKesson Speciality Health and North End Advisory. SH and TJB have patents relevant to manuscript.
Authorship
All named authors meet the International Committee of Medical Journal Editors (ICMJE) criteria for authorship for this article, take responsibility for the integrity of the work as a whole, and have given their approval for this version to be published.
Authors’ Contributions
All authors participated in the drafting, critical revision, and approval of the final version of the manuscript. SH led study design and conducted the bioinformatic analysis. SD conducted the statistical analysis. SH was primarily responsible for interpretation of the study results, with contributions from all authors.
Ethics Approval
Consent to Participate
All subjects provided written informed consent.
Acknowledgments
The authors thank all clinicians for their involvement and contribution to the study. The authors thank Kate Hendricks, MD, MPH&TM for many helpful editorial suggestions. Finally, the authors thank Sydney M Finegold, MD for mentorship that sparked the interest in the microbiome to many scientists, including authors of this paper.
Medical writing assistance was provided by Jordana Campbell, BSc, CMPP and Sonya Dave, PhD and was funded by ProgenaBiome.
Footnotes
Financial Information: No funding was received for this manuscript. Several authors were compensated or salaried by ProgenaBiome.
References



