
Abstract
The SARS-CoV-2 pandemic has resulted in widespread morbidity and mortality globally, but with widely variable outcomes. The development of severe disease and mortality is higher in older individuals, males and those with other co-morbidities, and may vary across ethnic groups. However, so far, no host genetic factor has been clearly associated with susceptibility and development of severe disease. To understand the impact of host genetics on expression of ACE2 (SARS-CoV-2 receptor) during RNA virus infection we performed a GWAS for ACE2 expression in HCV-infected liver tissue from 195 individuals. We discovered that polymorphisms in the host IFNL region which control expression of IFNL3 and IFNL4 modulate ACE2 expression. ACE2 expression was regulated additionally by age, with a subsidiary effect of co-morbidity. The IFNL locus controlled expression of a gene network incorporating many well-known interferon-stimulated genes which anti-correlated with ACE2 transcript levels. The same anti-correlation was found in the gastrointestinal tract, a site of SARS-CoV-2 replication where inflammation driven interferon-stimulated genes are negatively correlated with ACE2 expression. The interferon dependent regulation of ACE2 was identified in a murine model of SARS-CoV-1 suggesting conserved regulation of ACE2 across species. Polymorphisms in the IFNL region, as well as age, may impact not only on classical antiviral responses but also on ACE2 with potential consequences for clinical outcomes in distinct ethnic groups and with implications for therapeutic interventions.
Introduction
Severe acute respiratory syndrome–coronavirus 2 (SARS-CoV-2) which results in coronavirus disease 2019 (COVID-19) is a positive-stranded RNA virus that causes a severe respiratory syndrome in a subset of infected individuals and has led to widespread global mortality. The genome of SARS-CoV-2 shares about 80% sequence similarity with SARS-CoV and 96% sequence similarity with bat coronavirus Bat CoV RaTG131.
Entry of coronaviruses into susceptible cells depends on the binding of the spike (S) protein to a specific cell-surface protein and subsequent S protein priming by cellular proteases. Similar to SARS-CoV-1, infection by SARS-CoV-2 employs ACE2 as a receptor for cellular entry2. Viral entry also depends on TMPRSS2 protease activity, and cathepsin B/L activity may be able to substitute for TMPRSS22.
Epidemiological studies have indicated that the risk for serious disease and death from COVID-19 is higher in males, in older individuals and those with co-morbidities3–5. Host genetic variation is important in determining susceptibility and disease outcome for many infectious diseases6 and it is likely to be important in determining SARS-COV-2 susceptibility and development of severe disease after infection. Such insights would be important in understanding pathogenesis, repurposing antiviral drugs and vaccine development.
The earliest immune defence mechanism activated upon virus invasion is the innate immune system7. Virus-induced signalling through innate immune receptors prompts extensive changes in gene expression which are highly effective in resisting and controlling pathogens and subsequently prompt the activation of inflammatory and or antiviral immune effectors involved in pathogen clearance8. It has been shown that host genetics contributes to transcriptional heterogeneity in response to infections9, which underlies some of the differences in innate immune responses observed between individuals and the varying susceptibility to infection10. Therefore in the context of infectious diseases, it is of paramount importance to investigate infected tissue to understand how host genetics may impact on gene expression.
To understand the impact of host genetics on expression of ACE2 in the presence of viral infection, we first used HCV-infected liver biopsies from 195 individuals and performed host genome-wide genotyping and liver transcriptomics. Performing a genome-wide association study (GWAS) for ACE2 expression, we found that ACE2 expression is modulated by host genetic variation in the IFNL region on chromosome 19. We also observed that increase in age is significantly associated with increase in ACE2 expression and that expression of interferon pathway signalling genes are negatively correlated with ACE2 expression. This pattern is conserved across tissues, infections and species.
Results
We used genotyped autosomal SNPs in the host genome to undertake a GWAS, where the trait of interest was the expression of ACE2 in virus-infected liver, performing more than 300,000 association tests. We performed linear regression assuming an additive model, and adjusting for population structure by including the first five host genetic principal components (PCs) as covariates. There was no inflation in the association test statistics (Supplementary Figure 1). We used a false discovery rate (FDR) of 5% to decide on significance.
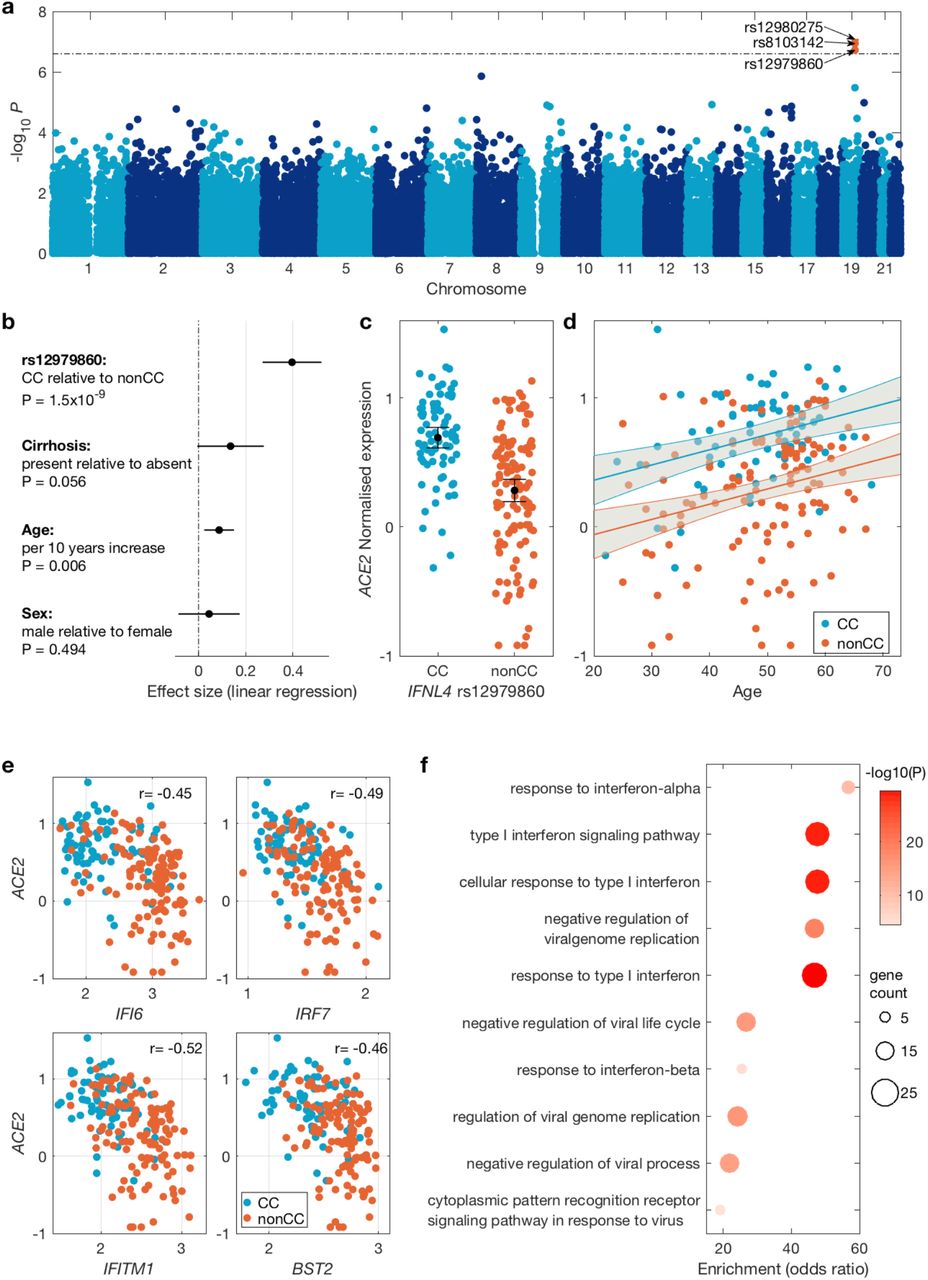
Across the human genome, the most significant associations were observed for three SNPs (all in linkage disequilibrium11 in European populations, Supplementary Figure 2 and 3) in the IFNL locus on chromosome 19 (Figure 1a and Supplementary Table 1, min p-value = 8.1×10−8 for SNP rs12980275).

(a) Manhattan plot of association between host genetic variation and ACE2 expression in virus infected liver biopsies using an additive model. The dashed line indicates 5% FDR level. Significant SNPs are coloured red and their ID is shown. (b) Forest plot of the effect sizes of SNP rs12979860 (dominant model), cirrhosis status, age and gender on ACE2 expression. The black circles indicate the point estimate and the black lines indicate their 95% confidence interval. (c) Distribution of ACE2 expression stratified by SNP rs12979860 genotypes (dominant model). Black circle shows the mean and the lines indicate its 95% confidence interval. (d) The relationship between ACE2 expression and age. The blue and red lines show the linear regression fit (for CC and nonCC genotypes respectively) and the gray area indicates their 95% confidence interval. (e) Expression of four representative interferon stimulated genes (ISG) and their observed negative correlation with ACE2 expression. The Pearson’s correlation coefficient is shown for each gene. (f) Gene Ontology gene set enrichment analysis among genes with significant negative correlation with ACE2 expression. Only the top ten enriched gene sets are shown, which are all ISG related pathways.
Host genetic variations in this region have previously been associated with HCV infection outcomes such as viral load, spontaneous clearance and treatment outcome, as well as viral evolution10,12,13. The causal variant is likely to be the dinucleotide exonic variant rs368234815 in IFNL414. This variant [ΔG > TT] causes a frameshift, abrogating production of functional IFN-λ4 protein. This variant is not directly typed on our genotyping array, however it is in high linkage disequilibrium with SNP rs12979860 [T > C] (r2 = 0.975 CEU population, 1000 Genomes dataset, rs12979860 T allele is in linkage with rs368234815 ΔG allele) which is an intronic SNP in the IFNL4 gene and was directly typed on our genotyping array and is one of the three SNPs significantly associated with ACE2 expression. The allele frequency varies substantially between populations globally, with the rs12979860 CC genotype (which protects against HCV) most enriched in East Asian populations and the non-CC genotypes (CT and TT) strongly enriched amongst those of African origin15 (Supplementary Figure 4).
To further understand the impact of polymorphisms in the host IFNL4 gene on ACE2 expression in presence of viral infection, we investigated the impact of IFNL4 SNP rs12979860 on expression of ACE2 in HCV-infected liver. We used linear regression using a dominant genetic model: CC vs CT and TT genotypes (i.e. those that do not produce IFN-λ4 protein and those that do) including age, gender and cirrhosis status as covariates (Figure 1b). We observed significantly higher expression of ACE2 (P=1.5×10−9) in individuals with CC vs. non-CC genotypes (Figure 1c). Additionally, we observed that ACE2 expression increased with age (P=0.006) in both CC and non-CC patients (Figure 1d). Patients with cirrhosis tend to have higher ACE2 expression although this association was marginally not significant (P=0.056, Figure 1b).
The IFNL4 gene itself is polymorphic and a common amino acid substitution (coded by the SNP rs117648444 [G > A]) in the IFN-λ4 protein, which changes a proline residue at position 70 (P70) to a serine residue (S70), reduces its antiviral activity in vitro16,17. Patients harbouring the impaired IFN-λ4-S70 variant display lower hepatic interferon-stimulated gene (ISG) expression levels, which is associated with increased HCV clearance following acute infection and a better response to IFN-based therapy, compared to patients carrying the more active IFN-λ4-P70 variant16. After imputing and phasing IFNL4 rs368234815 and rs117648444 we observed three haplotypes: TT/G (IFN-λ4-Null); ΔG/G (IFN-λ4-P70) and ΔG/A (IFN-λ4-S70). HCV-infected patients were classified into three groups according to their predicted ability to produce IFN-λ4 protein: (i) no IFN-λ4 (two allelic copies of IFN-λ4-Null, N = 69), (ii) IFN-λ4–S70 (two copies of IFN-λ4-S70 or one copy of IFN-λ4-S70 and one copy of IFN-λ4-Null, N = 21), and (iii) IFN-λ4-P70 (at least one copy of IFN-λ4-P70, N = 92). Analysis of the IFN-λ4 predicted patient groups revealed that IFN-λ4–S70 group had the expected impact on ACE2 expression i.e. lower levels of ACE2 expression relative to the IFN-λ4-Null group (P=0.022), and higher levels relative to the (more stimulatory) IFN-λ4-P70 group although the effect was marginally not significant, P=0.087, Supplementary Figure 5).
We also investigated the impact of the IFNL4 SNP rs12979860 genotypes on TMPRSS2, CTSB and CTSL genes which may also be needed for viral entry2. We observed that these three genes had much higher expression levels in the liver relative to ACE2, however their expression was not significantly associated with SNP rs12979860 genotypes (Supplementary Figure 6). Both CTSB and CTSL genes had significantly lower expression in patients with cirrhosis (PCTSB=1.2×10−5, PCTSL=8.8×10−8), while TMPRSS2 had higher level of expression in cirrhotic patients; however this was not significant (P=0.11, Supplementary Figure 7).
We also performed correlation analysis accounting for multiple testing to identify genes correlated with ACE2 in virus-infected livers. We observed large correlation coefficients (maximum of 0.51) and detected 591 genes significantly correlated with ACE2 expression at 1% FDR and with correlation coefficients of > 0.3 or < −0.3. Considering separately the genes that were positively correlated and those that were negatively correlated with ACE2 expression (Supplementary Tables 2 and 3), we performed a gene set enrichment analysis, observing that genes involved in type I interferon signalling pathways were enriched among genes negatively correlated with ACE2 expression (Figure 1e and 1f). These genes overlap strongly with those induced by IFNL7. We also observed that genes involved in extracellular structure organisation were enriched among genes positively correlated with ACE2 expression (Supplementary Figure 8).
To understand the impact of the IFNL locus on the overall gene expression in the virus-infected liver, we used our liver transcriptome data and tested for association between SNP rs12979860 and gene expression data using a dominant genetic model (CC vs. CT and TT genotypes). At 1% FDR, SNP rs12979860 was an eQTL for 583 genes. Genes involved in type I interferon signalling were enriched among genes that were upregulated in non-CC individuals relative to the CC individuals. Genes involved in B and T cell mediated immunity were enriched among genes upregulated in CC individuals relative to non-CC individuals (Supplementary Figures 9 and 10).
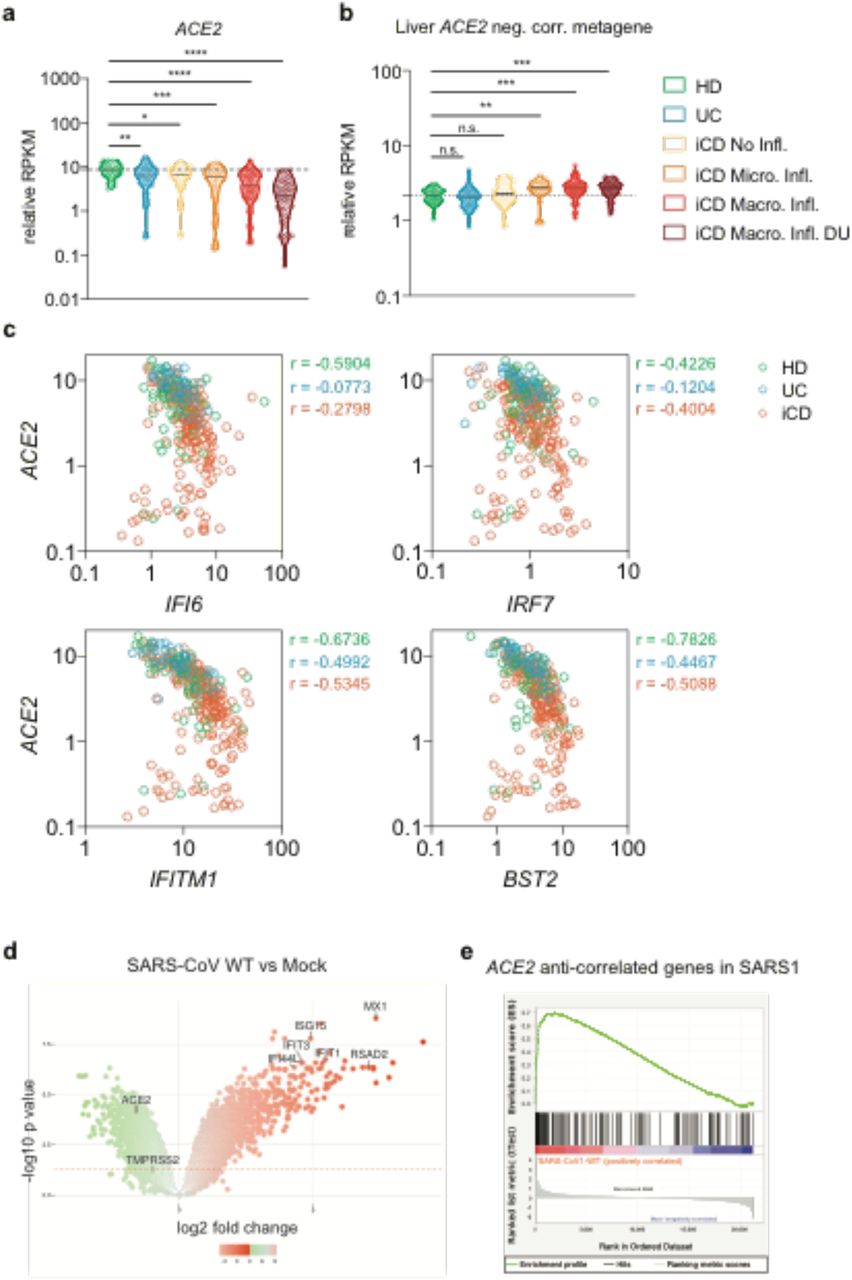
To further explore the anti-correlation of ISGs with ACE2 expression in a known site of SARS-CoV-2 replication, we explored the relationship between ACE2 and ISGs expression in the gastrointestinal (GI) tract, examining the levels of IFN-regulated genes in a gene expression study of terminal ileum biopsies in inflammatory bowel disease (IBD) in treatment-naive young donors (RISK cohort18). In intestinal biopsies, there was a striking decrease of ACE2 expression with increasing severity of inflammation that was independent of the abundance of transcriptional markers of epithelial identity19 (Figure 2a, Supplementary Figure 11a) and genes anti-correlated with ACE2 had increasing expression with rise in disease activity (Figures 2b and Supplementary Figure 11b). Genes associated with epithelial cell structure and function were enriched among genes that were positively correlated with ACE2 in both liver and intestine, while genes associated with type I interferon signalling pathways were enriched among genes that negatively correlated with ACE2 expression in both tissues (Figure 2c and Supplementary Figure 11c).

(a) ACE2 expression in terminal ileum biopsies transcriptomes of the RISK cohort grouped based on health state and histologic assessment of inflammation. (HD = Healthy Donor; UC = Ulcerative Colitis without ileal involvement; iCD = ileal Crohn’s Disease; Micro. Infl. = Microscopic Inflammation, Macro. Infl. = Macroscopic Inflammation; DU = Deep Ulcers). Data are shown as RPKM relative to the epithelial cell identity metagene (see methods) (b) Expression of liver ACE2 expression anti-correlated genes in the RISK cohort. Kruskal-Wallis test with multiple comparison correction controlling the FDR was used for association testing. (c) Expression and correlation of representative interferon stimulated genes (ISG) and ACE2 in the RISK cohort. Pearson’s correlation coefficients are shown. (d) Volcano plot of differential gene expression pattern induced by SARS-CoV1 infection in mouse model vs mock. Representative ISG genes and ACE2 are indicated. (e) GSEA plot of genes negatively correlated with ACE2 expression (enriched in ISGs) in the SARS-CoV-1 infected mouse model vs mock.
We repeated this analysis to define the impact of IFNL4 polymorphisms on gene expression in a second cohort of IBD patients enriched for those of African-American ethnicity20 (Supplementary Figure 11d). This analysis confirmed the clear anti-correlation of ACE2 expression with ISGs. Consistent with the absence of viral infection, there was no association seen between IFNL4 genotype and ACE2 expression in this IBD disease cohort (P=0.4).
Since the pattern of gene expression incorporating downregulation of ACE2 was consistent in two models of chronic infection and/or inflammation in different sites, we addressed whether a similar pattern of gene regulation was observed in lung tissue using data from mouse models of SARS infection21 (GSE59185). Indeed we observed in SARS-CoV-1 infected lung a similar enrichment of ACE2 regulating genes as observed in human liver. There was a strong correlation of gene regulation measured by GSEA analysis and furthermore we observed the same associated down-regulation of ACE2 in the presence of up-regulation of classical ISGs (Figure 2e and f).
Discussion
To understand the host genetic factors that drive ACE2 expression in the presence of RNA virus infection, we performed a genome-wide association analysis, for ACE2 expression in infected liver. Using infected tissue is important, since genetically driven differences in innate immune responses are only likely to be observed when innate immune responses are triggered. We observed that host genetic polymorphisms in the IFNL region modulate ACE2 expression in the presence of viral infection. The likely causal mechanism is the variant rs368234815 [ΔG > TT], which results in a frameshift and abrogates production of IFN-λ4 protein. Although our initial observation was made in patients with HCV infection and the liver tissue, given the robust maintenance of the transcriptional pattern in the GI tract and in murine models of SARS, it seems likely to be relevant to SARS-CoV-2 pathogenesis.
Interferon lambda receptor (IFNLR1) is largely restricted to tissues of epithelial origin22,23, therefore, IFN-λ proteins may have evolved specifically to protect the epithelium. Overall, INFL genes lead to a pattern of gene expression which is similar to type I interferon genes, but the time course and pattern of expression may vary7. This has been explored in HCV, where a slower, but sustained impact of IFNL signalling is seen24. In vitro studies have revealed that ISG expression and anti-viral activity induced by recombinant IFNL4 are comparable to that induced by IFNL325. Importantly, however, the tight regulation of IFNL426 means its ability to respond and induce a rapid antiviral state may be limited27 as seen both in vitro, and in vivo28. However, once established, the IFNL4 transcriptional module may also be highly sustained (as seen here and in other HCV cohorts16) and also noted elsewhere, e.g. after childbirth29.
In mice, the type III IFN response is restricted largely to mucosal epithelial tissues, with the lung epithelium responding to both type I and III IFNs30 and intestinal epithelial cells responding exclusively to type III IFNs. Among nonhematopoietic cells, epithelial cells are potent producers of type III IFNs. In mouse models, type III IFNs seem to be the primary type of IFN found in the bronchoalveolar lavage in response to influenza A virus infection and play a critical role in host defence31. Intriguingly, in humans, the IFNL4 polymorphism identified here is associated with the outcome of RNA virus respiratory tract infections in children, with the non-CC variant showing a poorer outcome32. The data from the GI tract indicate that this gene expression pattern is conserved amongst tissues, consistent with emerging data33. Inflammatory signals may act to sustain the triggering of ISGs and sustain downregulation of ACE2. Of note for inflammatory bowel disease, loss of ACE2 in the ileum impacts on secretion of antimicrobial peptides and the local microbiome34.
Downregulation of ACE2 itself may limit the ability of coronaviruses to enter cells, but may, if sustained, have impacts on inflammation. Indeed ACE2−/- mice suffer from enhanced disease following virus infection of the lung through an angiotensin-driven mechanism35. In other settings, the nonCC genotype may provide a more limited early response26 and lead to more sustained activation of the IFNL pathway29 than CC genotype (Supplementary Figure 12 and 13) although overall the issue of which genotype might be protective in COVID-19 remains open. We also note the impact of ageing, which blunts this response in both genotypes. The mechanism for this requires further study, as does the impact of gender. These data and model are also consistent with transient upregulation of ACE2 seen in early time points by IFNα in vitro36 but the full kinetics need further study in vitro and in vivo.
This study provides an orthogonal investigation of SARS-CoV-2 induction of ACE2. Although we did not study this directly in the respiratory tract, such studies should be urgently performed to confirm these data – ideally in epithelial tissue, where the model suggested above can be tested. Furthermore the overall impact of this polymorphism on the clinical course should be assessed, especially given the very variable distribution of IFNL4 alleles in different ethnic groups, which may in turn reflect selection earlier in human evolution10,15. Finally, the strong genetic data add weight to the idea of a careful exploration of IFNL pathways in therapy for SARS-CoV-237.
Data Availability
Information on access to individual-level genotype and sequence data is available here: http://www.stop-hcv.ox.ac.uk/data-access.
Methods
Boson cohort
Boson patient cohort
For this study, we used patient data from the BOSON cohort that has been described elsewhere38. All patients provided written informed consent before undertaking any study-related procedures. The BOSON study protocol was approved by each institution’s review board or ethics committee before study initiation. The study was conducted in accordance with the International Conference on Harmonisation Good Clinical Practice Guidelines and the Declaration of Helsinki (clinical trial registration number: NCT01962441).
RNA extraction, library prep, sequencing and mapping for the BOSON cohort
Liver biopsy samples were available for 198 patients. Total RNA was extracted from patient liver biopsies at baseline (pre-treatment) using RNeasy mini kits (Qiagen). Briefly, liver biopsy samples were mechanically disrupted in the presence of lysis buffer and homogenized using a QIAshredder. Tissue lysates were then centrifuged and clarified supernatants were transferred into new microcentrifuge tubes (pellets were discarded). Next, 1 volume of 70% ethanol was added to the lysates and samples were mixed by gentle vortexing. 700uL of sample was then transferred into RNeasy spin columns (with 2mL collection tubes) and centrifuged at 10000 rpm for 15 seconds. Column flow-through was discarded. DNase digestion was subsequently performed to eliminate any contamination from genomic DNA. 80uL of DNase I solution (10uL DNase I stock + 70uL Buffer RDD) was added directly to RNeasy spin columns and incubated at room temperature for 15 minutes. Following DNase incubation, the columns were washed with 350uL of Buffer RW1 and centrifuged at 10000 rpm for 15 seconds. Flow-through was discarded and 500uL of Buffer RPE was added to the spin columns. Columns were then centrifuged again at 10000 rpm for 15 seconds and flow-through was discarded. An additional 500uL of Buffer RPE was added to the spin columns and columns were centrifuged at 10000 rpm for 2 minutes. Finally, spin columns were transferred into new microcentrifuge tubes and 30uL of RNase-free water was added directly to the column membrane. Columns were then centrifuged at 10000 rpm for 1 minute to elute the RNA.
RNA yield was quantified using a NanoDrop spectrophotometer. Selected samples were also run on an Agilent TapeStation system to assess RNA quality and purity. Library preparation from purified RNA samples was performed using the Smart-Seq2 protocol39, used along with previously described indexing primers during amplification40.
High-throughput RNA sequencing of prepared libraries was performed on the Illumina HiSeq 4000 platform to 75bp PE at the Wellcome Center for Human Genetics (Oxford, UK). Reads were trimmed for Nextera, Smart-seq2 and Illumina adapter sequences using skewer-v0.1.12541. Trimmed read pairs were mapped to human genome GRCh37 using HISAT2 version 2.0.0-beta42. Uniquely mapped read pairs were counted using featureCounts43, subread-1.5.044, using exons annotated in ENSEMBL annotations, release 75. Mapping QC metrics were obtained using picard-tools-1.92 CollectRnaSeqMetrics.jar. Three samples were excluded after QC checks due to low sequencing depth which left 195 samples for analysis. Genes were filtered using the criteria of having a count per million of 1.25 in at least 10 samples to remove low expressed genes. Function cpm from edgeR45 version 3.20.9 was used to calculate the counts per million (CPM) values. Two samples had zero CPM values for ACE2 gene and they were set to the minimum ACE2 expression (in CPM) observed across all the samples. Log10 of CPM was used for all the analysis.
Host genotyping and imputation
Genomic DNA was extracted from buffy coat using Maxwell RSC Buffy Coat DNA Kit (Promega) as per the manufacturer’s protocol and quantified using Qubit (Thermofisher). DNA samples from patients were genotyped using the Affymetrix UK Biobank array, as described elsewhere12. Both liver RNA trascriptomic and human genome-wide SNP data were obtained on a total of 190 patients of mainly White self-reported ancestry infected with HCV subtype 3a. After quality control and filtering of the human genotype data, approximately 330,000 common SNPs with minor allele frequency greater than 5% were available for analysis. The Phasing and imputation was performed using SHAPEIT246 and IMPUTE247 version 2.3.1 using default settings and the 1000 Genomes Phase III dataset as a reference population48. After QC, the imputation data for 182 patients were of high quality. Imputation quality for samples included for rs117648444 and rs368234815 variants (information 0.974 and 0.994 respectively and certainty 0.995 and 0.997 respectively). All patients were also independently genotyped for SNP rs12979860 as described previously38.
Statistical analysis
To test for association between human SNPs and ACE2 expression (in log10(CPM)), we performed linear regression using PLINK49 version 1.9 using an additive genetic model adjusted for the human population structure (first five PCs). For 190 patients both host genome-wide genotyping data and liver RNAseq data was available. We used the qvalue package in R to calculate false discovery rate in this analysis and 5% FDR as significance threshold.
To test for association between ACE2 expression and host SNP rs12979860 (dominant genetic model (CC vs. CT and TT genotypes)), cirrhosis status, sex and age we used multivariate linear regression as implemented in R. To test for association between ACE2 expression and IFN-λ4 predicted patient groups (IFN-λ4-Null, IFN-λ4-S70 and IFN-λ4-P70), we used linear regression as implemented in R and added cirrhosis status, sex and age as covariates to the analysis.
Log(CPM) data as calculated by cpm function from edgeR package was used to calculate Pearson’s correlation coefficient against ACE2 expression. The qvalue package was used to calculate false discovery rate. To filter out significant genes we used FDR of 1% and correlation coefficient of >0.3 or <−0.3 for positively and negatively correlated genes. To test for enrichment we used enrichGO function from the clusterProfiler package50. We only investigated gene sets in “biological process” GO hierarchy.
To test for association between SNP rs12979860 genotypes (CC vs. CT and TT) and expression of all genes, we used LIMMA package with voom transformation51. Cirrhosis status, sex, age, race and batch number were included as covariate to account for possible confounders. The gene set enrichment analysis was performed using ernichGO function from the clusterProfiler package.
RISK cohort
RNA isolation cDNA synthesis
Cell lysates were homogenized with a QIAshredder column (Qiagen, Crawley, UK) and RNA extracted with the RNEasy Mini Kit (Qiagen, Crawley, UK) following manufacturers instruction. cDNA was reverse-transcribed from template RNA either using a two-step reverse transcription using AppScript cDNA synthesis kit (Appleton Woods).
RISK cohort classification and data analysis
The RISK study is an observational prospective cohort study with the aim to identify risk factors that predict complicated course in pediatric patients with Crohn’s disease52. The RISK study recruited treatment-naive patients with a suspected diagnosis of Crohn’s disease. The Paris modification of the Montreal classification were used to classify patients according to disease behaviour (non-complicated B1 disease (non-stricturing, non-penetrating disease); complicated disease, composed of B2 (stricturing) and/or B3 (penetrating) behaviour) as well as disease location (L1, ileal only, L2, colonic only, L3, ileocolonic and L4, upper gastrointestinal tract). 322 samples were investigated with ileal RNA-seq. Individuals without ileal inflammation were classified as non-IBD controls. Patients with Crohn’s disease were followed over a period of 3 years. Patients were largely of European (85.7%) and African (4.1%) ancestry. RPKM expression values for the RISK cohort52 were retrieved from GEO (GSE57945). The dataset was filtered to (n=19,556) genes that had an expression value ≥ 0.1 in >10% of the patients.
To account for the potential loss of epithelial cells contribution to gene expression a metagene score was generated based on the average expression of epithelial identity genes19. RPKM data were transformed and presented as: RPKM+1/epithelial cell metagene. For the intersection of ACE2 correlated gene expression, genes were ranked based on their pearson correlation coefficient to ACE2 for each patient subgroup. Intersected lists of ACE2 expression positively (pearson correlation coefficient > 0.5) and negatively (pearson correlation coefficient < −0.5) correlated genes were extracted (positive correlation: n = 2067; negative correlation: n = 2264). Liver ACE2 expression and RISK ACE2 expression correlated and anti-correlated gene sets were intersected based on Entrez gene identifiers using Cytoscape (version 3.7.1) and visualized using the Cytoscape Venn and Euler Diagrams (Version 1.0.3) plugin (http://apps.cytoscape.org/apps/vennandeulerdiagrams). Functionally grouped networks of terms and pathways were analysed using the Cytoscape (version 3.7.1) ClueGO (version 2.5.6) and CluePedia (version 1.5.6) plug-in53. The analysis was performed by accessing the Gene Ontology Annotation (GOA) Database for Biologic processes, Cellular components, Immune system processes and Molecular function, the Reactome pathways database (https://reactome.org/) and the KEGG database (https://www.genome.jp/kegg/pathway.html). Only pathways with an adjusted enrichment p-value ≤ 0.05 were considered (Two-sided hypergeometric test, Bonferroni step down p-value correction). GO terms were grouped based on the highest significance when more than 50% of genes or terms were shared. The filtered RISK gene expression data (n=19,556; expression value ≥ 0.1 in >10% of the patients) served as reference gene set.
Resources for statistical analysis and data visualization:
Prism version 8.0 (GraphPad Software)
Excel for Mac Version 15.32 (Microsoft)
Cytoscape 3.7.1 (https://cytoscape.org/)
Cytoscape 3.7.1 plugin ClueGO (Version 2.5.6)
Cytoscape 3.7.1 plugin CluePedia (version 1.5.6)
Cytoscape 3.7.1 plugin Venn and Eluler Digrams (Version 1.0.3)
R (Version 3.6.1)
RStudio (Version 1.2.5001)
Specific statistical tests applied in this study are described in the respective figure legends. The level of statistically significant difference was defined as p ≤ 0.05.
GENESIS cohort
Ileal biopsies from GENESIS cohort
GENESIS is funded by the National Institute of Diabetes and Digestive and Kidney Diseases and managed by Emory University for the recruitment of self-identified African American subjects with IBD54. We used a subset of 195 GENESIS cohort subjects with ileal transcriptomic profiles as an additional replication cohort to test for anti-correlation of ACE2 expression with IFN gene expression. Pearson correlation tests between normalized expression values for ACE2 and four IFN genes confirmed that this pattern of anti-correlation is also observable in a cohort enriched for African American ancestry. This dataset includes 158 IBD patients along with 37 controls. Subjects with ileal inflammation were included as IBD, while non-IBD controls did not have ileal inflammation. This dataset is enriched for African American ancestry (70%), and gender was equally distributed. Full descriptions of age, gender, race, disease status and other phenotypic information are available in a prior publication20. Additionally, ileal transcriptomic profiles sequenced on the NextSeq 550 platform are available in the GEO repository (GSE57945) for all subjects.
RNA pre-processing and data analysis for SARS mouse model
RNA-Seq read counts from each samples were aligned to human reference hg38 using STAR55 and HISAT56 alignment algorithms and gene read counts were generated for each mapped sample using featureCounts program43. Low expressed features were filtered out prior logCPM transform of the read counts matrix, normalization and further gene filtering by expression variability (IQR > 0.75) and annotation were applied retaining 5817 genes to following analysis. EdgeR57 and limma51 packaged were used to perform differential gene expression analysis of in-home generated and public available data. Gene set enrichment analysis (GSEA58) was employed to measure co-expression of our lists of gene correlations and differentially expression analysis in public available data from GEO.
Acknowledgements
The authors would like to thank Gilead Sciences for the provision of samples and data from the BOSON clinical study for use in these analyses. The authors would also like to thank HCV Research UK (funded by the Medical Research Foundation) for their assistance in handling and coordinating the release of samples for these analyses. This work was funded by a grant from the Medical Research Council (MR/K01532X/1 – STOP-HCV Consortium). The work was supported by Core funding to the Wellcome Centre for Human Genetics provided by the Wellcome Trust (203141/Z/16/Z). This work was also supported by the Medical Research Council [grant number MC_PC_19059] and a strategic award from the Wellcome Trust (211276/Z/18/Z – WSSS). PK, is supported as a Wellcome Trust Senior Investigator (WT 109965MA) and an NIHR Senior Investigator. PK is affiliated to the National Institute for Health Research Health Protection Research Unit (NIHR HPRU) in Emerging and Zoonotic Infections at University of Liverpool in partnership with Public Health England (PHE), in collaboration with Liverpool School of Tropical Medicine and the University of Oxford. EB was funded by the Medical Research Council UK, the Oxford NIHR Biomedical Research Centre and is an NIHR Senior Investigator. The work was also supported by the NIHR Biomedical Research Centre, Oxford. The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR, the Department of Health or Public Health England. The authors thank Jan Rehwinkel for his contributions to the manuscript preparation and David Klenerman (University of Cambridge) for his contribution to the high throughput sequencing platform that underpins this work.




{kind=link}
{kind=link}