
ABSTRACT
Background Limited data are available on the long-term clinical and immunologic consequences of SARS-CoV-2 infection in people with HIV (PWH).
Methods We measured SARS-CoV-2 specific humoral and cellular immune responses in people with and without HIV recovering from COVID-19 (n=39 and n=43, respectively) using binding antibody, surrogate virus neutralization, intracellular cytokine staining, and inflammatory marker assays. We identified individuals experiencing symptomatic post-acute sequelae of SARS-CoV-2 infection (PASC) and evaluated immunologic parameters. We used linear regression and generalized linear models to examine differences by HIV status in the magnitude of inflammatory and virus-specific antibody and T cell responses, as well as differences in the prevalence of PASC.
Results Among PWH, we found broadly similar SARS-CoV-2-specific antibody and T cell immune responses as compared with a well-matched group of HIV-negative individuals. PWH had 70% lower relative levels of SARS-CoV-2 specific memory CD8+ T cells (p=0.007) and 53% higher relative levels of PD-1+ SARS-CoV-2 specific CD4+ T cells (p=0.007). Higher CD4/CD8 ratio was associated with lower PD-1 expression on SARS-CoV-2 specific CD8+ T cells (0.34-fold effect, p=0.02). HIV status was strongly associated with PASC (odds ratio 4.01, p=0.008), and the proportion of PD-1+ CD4+ T cells and levels of certain inflammatory markers (IL-6, TNF-alpha, and IP-10) were associated with persistent symptoms.
Conclusions We identified potentially important differences in SARS-CoV-2-specific CD4+ and CD8+ T cells that might have implications for long-term immunity conferred by natural infection. HIV status strongly predicted the presence of PASC. Larger and more detailed studies of PASC in PWH are urgently needed.
BACKGROUND
As the SARS-CoV-2 pandemic continues, its intersection with the ongoing HIV pandemic has gained increased attention [1]. While studies conducted early in the course of the pandemic did not show differences in outcomes of acute COVID-19 associated with HIV status [2–4], larger and more carefully designed studies support a growing consensus that people with HIV (PWH) are at higher risk for adverse outcomes [5–9]. In addition, the SARS-CoV-2 pandemic has caused massive disruption to systems of HIV care, potentially affecting access to care and antiretroviral therapy (ART) [10–13]. Data on SARS-CoV-2-specific adaptive immune responses in PWH compared with HIV-negative individuals remain sparse, with one recent study showing less robust immune responses among PWH [14] but another [15] suggesting similar responses. Furthermore, there is growing recognition of the clinical burden of post-acute sequelae of SARS-CoV-2 infection (PASC, including “long COVID”) [16], but this condition remains poorly understood, especially in PWH.
While the mechanisms underlying PASC have yet to be fully delineated, early studies have suggested that a combination of factors might contribute. These include innate and adaptive immune dysregulation [17–20], autoimmunity [21–23], human herpes virus reactivation [24], and microvascular dysfunction [25,26], among others [24]. Certain sociodemographic factors (e.g., income status, access to care) [27,28] and a greater number and/or certain types of medical comorbidities (e.g., obesity, diabetes mellitus, pre-existing cardiovascular or neurologic disease) [27,29,30] may also increase the likelihood of an individual developing PASC. Higher prevalence of such factors among PWH [31,32], along with differences in immune responses to SARS-CoV-2 [14,15] and chronic inflammation and immune dysregulation even in the presence of antiretroviral therapy [33– 39], may make PWH selectively vulnerable to developing persistent COVID-19-attributed symptoms following SARS-CoV-2 infection. For these reasons, examination of PASC in PWH is urgently needed [40].
In this study, we sought to characterize SARS-CoV-2 specific immune responses and the presence of persistent symptoms in a cohort of PWH with a history of COVID-19 prior to the availability of vaccination in comparison to a similar group of HIV-negative individuals recovering from COVID-19. We hypothesized that PASC would be more prevalent among PWH and that this would be driven by reduced SARS-CoV-2 specific immune responses and increased indices of systemic inflammation among this population.
METHODS
Informed consent
The study was approved by the Institutional Review Board at the University of California, San Francisco (UCSF). All participants provided written informed consent.
Participants
Volunteers were enrolled into the Long-term Impact of Infection with Novel Coronavirus (LIINC) COVID-19 recovery cohort at UCSF (NCT 04362150). Study procedures are described in detail elsewhere [41,42]. Briefly, participants with a prior positive nucleic acid amplification test confirming SARS-CoV-2 infection were enrolled >=21 days following symptom onset. Recruitment occurred through a combination of self-and clinician referrals; all PWH testing positive for SARS-CoV-2 at two UCSF-based HIV clinics were notified of the study and invited to participate. At each study visit, participants completed a structured interview with a research coordinator in which they provided clinical data regarding prior and current COVID-19 attributed symptoms, medical and HIV history, and quality of life.
For the current study, we selected all PWH who enrolled prior to the receipt of a SARS-CoV-2 vaccine (n=39) and compared them with HIV-negative individuals (n=43) with a similar distribution of key variables demonstrated to be important in determination of the SARS-CoV-2 immune responses in our cohort [17,43–45]: age, sex, hospitalization during COVID-19, and time since infection. We assessed their immunologic parameters and symptoms at the time point closest to 16 weeks post-infection (median 112 days [IQR: 91-129]).
Clinical assessment of PASC
Our methodology for assessing PASC in the cohort is described in detail elsewhere [42]. At each study visit, we tabulate responses to questions regarding the presence of 32 somatic symptoms derived from the U.S. Centers for Disease Control (CDC) list of COVID-19 symptoms [46] and the Patient Health Questionnaire Somatic Symptom Scale [47]. Importantly, symptoms are only considered as related to COVID-19 if they are new or worsened since the time of initial SARS-CoV-2 infection; stable, chronic symptoms that pre-dated SARS-CoV-2 infection are not included in the PASC definition.
For the primary analysis, we defined PASC as any COVID-19 attributed symptom that was present during a study visit >6 weeks following SARS-CoV-2 infection; this definition was selected because it is consistent with prior work and consensus definitions [48,49]. We conducted further sensitivity analyses using a stricter definition of PASC as 3 or more symptoms at this timepoint, comparing this group to those with fewer than 3 symptoms and to those with no symptoms.
Biospecimen isolation
Peripheral blood mononuclear cells (PBMCs), plasma and serum were obtained from participants and cryopreserved prior to testing. PBMCs were isolated using Ficol-Paque in SeptMate Tubes. PBMCs were frozen in heat inactivated FBS and 10% dimethyl sulfoxide (DMSO) and stored in liquid nitrogen.
Antibody assays
Virus-specific antibody responses were measured the Pylon COVID-19 total antibody assay (ET Health) and a validated surrogate virus neutralization test (sVNT) [50]. The sVNT measures competitive inhibition of the interaction of the original SARS-CoV-2 RBD and angiotensin-converting enzyme 2 (ACE-2). The lower limit of detection was a sVNT reciprocal titer <10 and anti-RBD IgG <10 relative fluorescence units.
T cell responses by Intracellular Cytokine Staining (ICS)
We performed ICS as previously described [44] with the following modifications. After thawing, PBMCs were rested overnight at 37°C at a concentration of 1×106 PBMC per well in RPMI plus 10% FBS, penicillin, streptomycin, and L-glutamine (R10). The following day, PBMCs were cultured for 18 hours at 37°C in 96-well U-bottom plates at 1×106 PBMC per well in R10 in the presence of either SARS-CoV-2 peptide megapools (described in [51]; [1 µg/ml/peptide]), or 0.25-1×106 PBMC per well with CD3/CD28 beads (Immunocult, STEMCELL Technologies) as a positive control or equimolar DMSO as negative control. Two previously-described peptide megapools containing SARS-CoV-2-derived peptides experimentally determined to be recognized by CD4+ or CD8+ T cells were used for peptide stimulation (CD4-E with 280 and CD8-E with 454 T cell epitopes) [51]. All conditions were in the presence of brefeldin A (Thermo Fisher), monensin (Thermo Fisher) and anti-CD107a. After an 18-hour incubation, cells were washed and surface markers were stained for 15 min at room temp in the dark. Following surface staining, cells were washed twice with PBS and then fixed/permeabilized (BD Cytofix/Cytoperm) for 20 min at 4°C in the dark. Cells were then washed twice with perm wash buffer (BD Perm/Wash) and stained with intracellular antibodies for 30min at 4°C in the dark. All samples were acquired on a BD LSR-II analyzer and analyzed with FlowJo X software. The flow cytometric gating strategy is shown in Supplementary Figure 1.
Markers of inflammation
In a subset of participants with additional specimens available, the fully automated HD-X Simoa platform was used to measure biomarkers in blood plasma including monocyte chemoattractant protein 1 (MCP-1), Cytokine 3-PlexA (IL-6, IL-10, TNFα), interferon gamma-induced protein-10 (IP-10), and interferon-gamma (IFNγ). Samples were assayed blinded with respect to associated patient and clinical information. Assay performance was consistent with the manufacturer’s specifications.
Statistical methods
We used descriptive statistics to characterize the cohort. For the comparison of humoral responses, we first log-transformed sVNT and total antibody values as relative differences are more clinically meaningful than absolute differences, and to satisfy assumptions of a normal distribution. We then used linear regression models to examine differences in magnitude of humoral models by HIV status, adjusting for days since onset of known SARS-CoV-2 infection, age, sex at birth, history of hospitalization for SARS-CoV-2 infection, and the primary indicator for the analysis, HIV status. Differences and 95% confidence intervals (CI) are presented as fold-changes in the geometric mean. Among PWH, CD4/CD8 ratio was then added to the model as an additional covariate to examine if CD4/CD8 ratio was associated with magnitude of responses among PWH. To examine percent differences in cellular immune responses by HIV status, we used generalized linear models from the binomial distribution with bootstrapped standard errors and adjusted for the same potential confounders as above. To examine differences in relative mean inflammatory markers by HIV-status, we fit linear regression models following log-transformation of the individual inflammatory markers. To examine differences in the experience of PASC by HIV status, we used logistic regression adjusting for the same potential confounders.
RESULTS
Participants
All participants were diagnosed with COVID-19 by nucleic acid amplification (NAA) testing between March and December 2020 (pre-Delta and Omicron) and none had received a SARS-CoV-2 vaccine prior to specimen collection. The groups were similar in terms of age, sex, and race/ethnicity (Table 1). The vast majority of PWH in the study were men, reflecting the demographics of the San Francisco Bay Area HIV epidemic [52]. While most participants were generally healthy, those with HIV more commonly reported concurrent medical comorbidities including heart and lung disease. The majority of participants were managed as outpatients during their acute COVID-19 illness. Among the minority who were hospitalized, all required supplemental oxygenation and only one participant in each group required mechanical ventilation. No participant reported receiving SARS-CoV-2-specific treatments during their acute illness.
Values reported as median (IQR) unless otherwise specified. IQR, interquartile range. Note: Autoimmune disease reported as one case of hypothyroidism and one case of Raynaud’s disease. Cancer reported as prostate cancer controlled with hormone therapy, ocular melanoma, treated Kaposi sarcoma, and resected renal cancer. Plasma HIV RNA values were >50 in two participants (87 copies/mL and 28,118 copies/mL). All hospitalized participants in each group required oxygen support and one hospitalized participant in each group required mechanical ventilation. None reported receiving SARS-CoV-2-targeted therapy or steroids during the hospitalization.
The median time between COVID-19 symptom onset and research assessment was 117 (85-128) days for PWH and 111 (94-131) days for HIV-negative individuals (p=0.58; Table 2). Initial illness severity was similar between both groups (13% vs 17% hospitalized, p=0.41) Participants reported a wide array of symptoms attributed to COVID-19 at the time of PASC assessment (Table 2). There were similarities in post-acute symptoms reported among both groups, but a larger proportion of PWH reported certain symptoms such as fatigue, gastrointestinal and certain neurocognitive symptoms, and issues with sleep.
Values reported as n (%). Participants were systematically asked about 32 individual symptoms at the late follow-up visit, which took place a median of 124 days from initial COVID-19 symptom onset.
SARS-CoV-2-specific antibody responses
Antibody responses were similar between people with and without HIV infection. In models incorporating HIV status, days since infection, age, sex, and prior hospitalization during SARS-CoV-2 infection (yes vs. no), HIV status was not a predictor of humoral responses as measured by SARS-CoV-2-specific antibody binding (1.31-fold higher; 95%CI: 0.70-2.46; p=0.40; Fig. 1a) and surrogate virus neutralization testing (1.01-fold higher; 95% CI: 0.63-1.63; p=0.95; Fig. 1b). Hospitalization during acute COVID-19 was associated with 4.29-fold higher geometric mean titers of binding antibodies (95% CI:1.74-10.57; p=0.002) and 2.57-fold higher titers of surrogate viral neutralization testing (95% CI: 1.33-4.98; p=0.005).

A. Total SARS-CoV-2 antibodies. B. Surrogate viral neutralization titers. C. SARS-CoV-2 specific CD4+ and CD8+ T cells. D. PD1 expression on SARS-CoV-2 specific CD4+ and CD8+ T cells. Bars represent median and interquartile ranges.
SARS-CoV-2 specific T cell responses
We measured SARS-CoV-2-specific memory T cells by intracellular cytokine staining (ICS) after stimulation with peptide megapools containing predicated SARS-CoV-2 CD4+ (CD4-E) or CD8+ (CD8-E) T cell epitopes, as described [51]. In models incorporating days since infection, age, sex, and prior hospitalization during SARS-CoV-2 infection, HIV status was not a predictor of the magnitude of interferon gamma (IFN□)-producing SARS-CoV-2-specific memory CD4+ T cell responses (1.13-fold higher; 95%CI: 0.76-1.71; p=0.53; median: 0.070% vs. 0.068%; Fig. 1c). However, PWH had 70% lower relative levels of SARS-CoV-2-specific memory CD8+ T cells (0.30-fold; 95% CI: 0.12-0.72; p=0.007; median: 0.034% vs. 0.016%; Fig. 1c.).
PWH exhibited higher levels of PD-1 expression on SARS-CoV-2-specific memory CD4+ T cells in adjusted regression analyses (1.53-fold higher; 95% CI: 1.12-2.10; p=0.007; median: 57.1% vs. 65.0%; Fig. 1d;), but no significant differences in PD-1 expression on SARS-CoV-2 specific CD8+ T cells (1.21-fold higher; 95% CI: 0.75-1.94; p=0.44; median: 24.1% vs. 25.0%; Fig. 1d).
Effect of CD4/CD8 ratio
The ratio of CD4+ to CD8+ T cells in the peripheral blood is a marker of general immune health in people with HIV [53]. Because a low CD4/CD8 ratio has been associated with reduced responses to vaccination against yellow fever in PWH [54], we next examined whether the CD4/CD8 ratio was associated with adaptive immune responses after infection with SARS-CoV-2 among PWH using adjusted regression modeling as above with the addition of CD4/CD8 ratio as a covariate. The ratio was not predictive of binding antibody levels (p=0.30) or surrogate virus neutralization (p=0.61). Higher ratios were associated with 67% lower overall frequency of SARS-CoV-2 specific CD4+ T cells (0.33-fold; 95% CI: 0.19-0.97; p=0.04) and 36% lower SARS-CoV-2 specific PD-1 expression (0.64-fold 95% CI:0.57-0.97; p=0.03). Notably, higher CD4/CD8 ratios were also associated with 66% lower PD-1 expression on SARS-CoV-2 specific CD8+ T cells (0.34-fold 95% CI: 0.13-0.87; p=0.02). There was a nonsignificant trend toward a similar finding among HIV-negative individuals (0.70-fold; 95% CI: 0.47-1.04; p=0.08).
Relationship between antibody and T cell immune responses
Similar to our prior work [43], we observed strong correlations between the binding (r=0.33, p=0.008) and surrogate viral neutralization response (r=0.33, p=0.007) and between binding and surrogate viral neutralization responses and SARS-CoV-2-specific CD4+ T cells (r=0.41, p<0.001 and r=0.42, p<0.001, respectively).
Markers of systemic inflammation
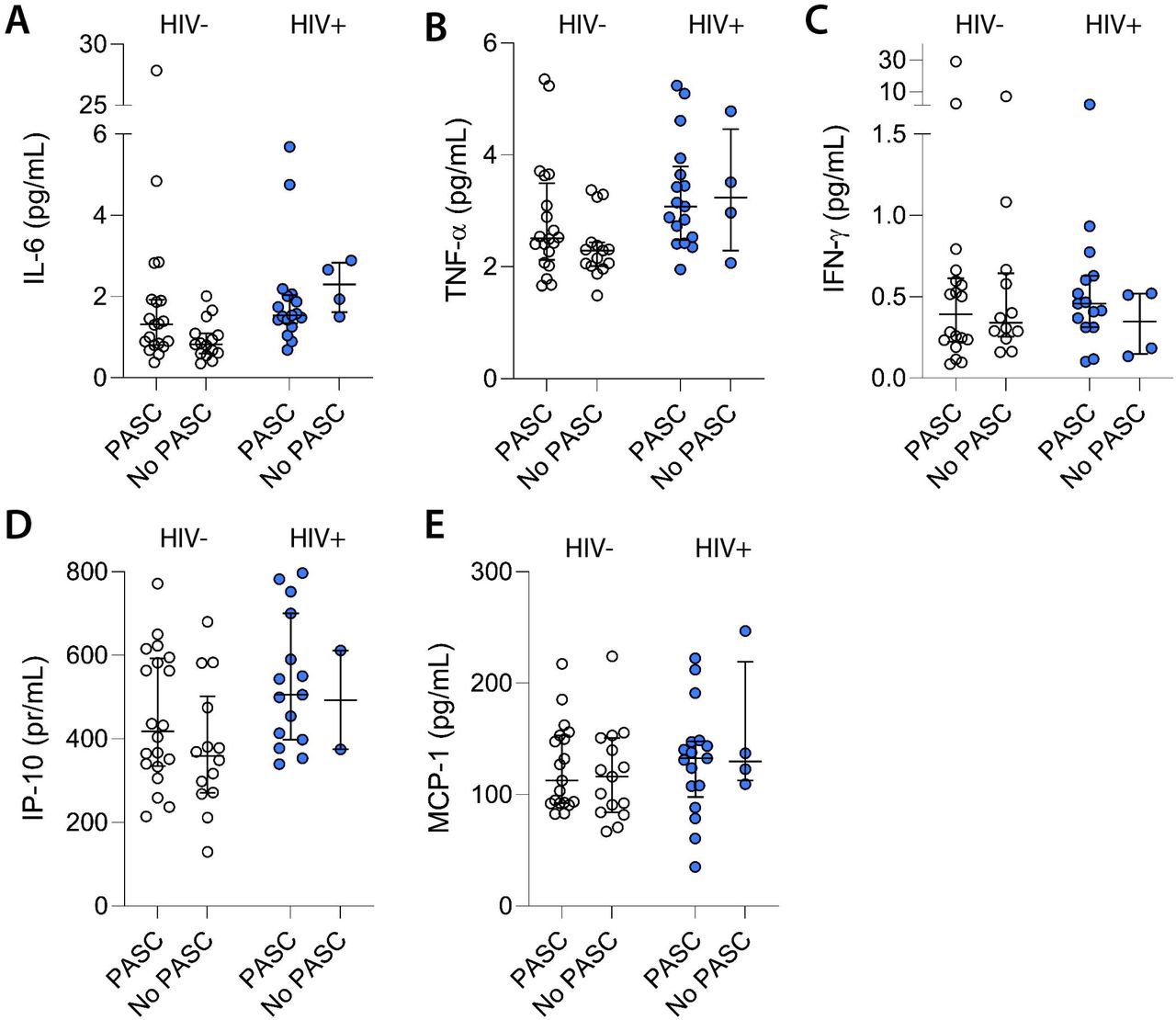
PWH had higher levels of plasma markers of inflammation compared with HIV-negative comparators. Among PWH, mean IL-6 levels were 1.55-fold higher (95% CI: 1.06-2.26; p=0.02), mean IP-10 levels were 1.31-fold higher (95% CI: 1.06-1.62; p=0.01), and TNFα levels were 1.26-fold higher (95% CI: 1.08-1.47; p=0.003) than in HIV-negative comparators.
Post-acute sequelae
We found HIV status to be highly associated with the presence of PASC. In the primary analysis defining PASC as any COVID-19 attributed symptom that was new or worsened since the time of SARS-CoV-2 infection and present at the time of the visit, PWH had 4.01-fold higher odds of reporting PASC (95% CI: 1.45-11.1; p=0.008), in a model controlling for time since infection, hospitalization, and age. This relationship was maintained when defining PASC as 3 or more symptoms in comparison to fewer than 3 symptoms (adjusted odds ratio (AOR) 2.72; 1.08-6.88; p=0.03). PWH reported more somatic symptoms overall (median 3 [IQR 1-6] versus median 1 [IQR 0-5], p=0.02), and in a model including time since infection, hospitalization, sex, and age, those with HIV had a 1.91-fold higher number of individual PASC symptoms than those without HIV (p=0.02).
Antibody and T cell immune responses did not correlate with PASC within the cohort (Fig. 2a-d). However, in models adjusting for HIV status, higher PD-1 expression on total non-naive CD4+ T cells, but not CD8+ T cells, was independently predictive of PASC when controlling for HIV-status (Fig. 2e-f). Each 10% decrease in PD-1 expression was associated with 2.02 fold-higher odds of PASC (AOR 2.02; 95% CI: 1.03-3.96; p=0.04).

A. Total SARS-CoV-2 antibodies. B. Surrogate viral neutralization titers. C. SARS-CoV-2 specific CD4+ T cells. D. SARS-CoV-2 specific CD8+ T cells. E. PD1 expression on bulk CD4+ T cells. F. PD1 expression on bulk CD8+ T cells. Bars represent median and interquartile ranges.
We found that some inflammatory markers were associated with increased odds of PASC (Fig. 3a-e). After adjusting for HIV status, the odds of PASC increased 1.18-fold for each 10% increase in IP-10 (AOR 1.18; 95% CI:1.01-1.38; p=0.04); and 1.10-fold for each 10% increase in IL-6 (AOR 1.10; 95% CI: 1.01-1.21; p=0.04); while there was a trend in increased PASC with higher TNFα levels (AOR 1.19; 95% CI: 0.98-1.46; p=0.08). Other plasma markers of inflammation were not associated with experience of PASC, including IL-10 (p=0.61), interferon gamma (p=0.73), and MCP-1 (p=0.93). Among PWH, there were increased odds of PASC with each 10% increase in IP-10 levels (AOR 1.06; 95% CI: 1.00-1.11; p=0.05), and a trend for increased PASC with higher TNFα levels (AOR 1.20; 95% CI: 0.97-1.49; p=0.09), but not IL-6 (p=0.64). This analysis was limited by the relatively small number of individuals with HIV who reported full recovery.
{kind=link}
{kind=link}
{kind=link}
A. Interleukin-6 levels. B. TNF-alpha levels. C. Interferon-gamma levels. D. IP-10 levels. E. MCP-1 levels. Bars represent median and interquartile ranges.
DISCUSSION
We found that post-acute COVID-19-attributed symptoms were common in this cohort of mostly ART-treated PWH recovering from SARS-CoV-2 infection. HIV status was strongly associated with PASC, raising concerns that this condition might be common among PWH recovering from COVID-19. Among those living with HIV, we observed broadly similar SARS-CoV-2-specific antibody and T cell immune responses as compared with a well-matched group of HIV-negative individuals, and higher levels of markers of inflammation that were also associated with PASC. Finally, we observed potentially important differences in SARS-CoV-2 specific CD4+ and CD8+ T cells that might have implications for long-term immunity conferred by natural infection. This analysis adds to the limited published data examining SARS-CoV-2-specific immune responses in PWH and underscores the need for larger and more detailed studies of PASC in this population.
An unknown proportion of people experience PASC following acute COVID-19, and there are currently massive efforts underway to understand the pathophysiology of this condition. There are limited data on PASC among PWH [55], however. One single-center study suggested that acute illness severity in PWH was significantly associated with PASC with 44% of participants experiencing at least one symptom 30 days after recovery [40], however it did not find an association between clinical parameters such as CD4+ T cell count, viral load, demographics or comorbidities and the presence of PASC, and did not compare PWH with HIV-negative individuals. Another retrospective study suggested that HIV was one factor that was associated with higher risk of PASC approximately one year after COVID-19 diagnosis among those requiring emergency department or hospital-based care [56], but the analysis was limited to 10 PWH and did not include biological measurements. While our cohort is not representative of all individuals with SARS-CoV-2 infection and cannot be utilized to estimate the population-level prevalence of PASC in the overall population or among PWH, the observation that persistent SARS-CoV-2-attributed symptoms were highly prevalent in PWH and that the adjusted odds of PASC were nearly four-fold as high as in a relatively well-matched HIV-negative comparator group was striking. Further description of this condition among PWH is urgently needed, including both detailed cohort descriptions and large-scale epidemiologic studies in which HIV can be examined as a predictor of PASC.
There are biological reasons why PWH might be particularly predisposed to developing PASC. One leading hypothesis is that PASC is driven, at least in part, by residual or ongoing inflammation following SARS-CoV-2 infection [17–20]. Even with suppressive ART, HIV is a chronic inflammatory condition that can be associated with complications of persistent immune activation [33–39]. If a higher prevalence of PASC among PWH recovering from COVID-19 is confirmed in larger epidemiologic studies, one possible explanation could be that this is driven in part by immune dysregulation from SARS-CoV-2 infection compounded upon higher baseline inflammation from HIV infection, which in turn could drive organ system dysfunction and the experience of somatic symptoms. In addition to elevated inflammatory responses, additional factors that could potentially predispose PWH to PASC include an increased frequency of autoimmune phenomena [57–59], localized tissue inflammation related to mucosal and endothelial injury [31,32], and other syndemic comorbidities including substance use [60,61] and metabolic disorders [62–64]. Furthermore, reactivation of latent human herpes viruses, such as Epstein Barr Virus or Cytomegalovirus (CMV) during acute SARS-CoV-2 infection may further perturb immune dysregulation and promote inflammation. PWH tend to have higher rates of CMV infection than HIV-negative individuals and this is thought to drive, in part, chronic inflammation in treated HIV and may lead to impaired CD8+ T cell function. Regardless of the mechanism, our observation is of importance because it suggests that PASC may be especially common in PWH and emphasizes the urgent need for larger studies of PASC in this population, which are underway [65].
Data on SARS-CoV-2-specific adaptive immune responses in PWH compared with HIV-negative individuals remain limited. Given the clear association between the presence of potent, durable SARS-CoV-2-specific immune responses and protection from disease upon re-exposure, it is critical to understand how HIV infection modulates protective antiviral immunity in PWH. Furthermore, there is evidence that SARS-CoV-2 can cause chronic infection in certain immunocompromised individuals [66], including those with advanced HIV infection [67], and result in viral evolution and the emergence of novel variants of concern. Our findings of similar humoral immune responses between well-matched groups of individuals with and without HIV infection sampled at similar time points after infection add to a limited literature on adaptive immunity to natural SARS-CoV-2 infection in PWH. Since the beginning of the pandemic, there has been concern that PWH might be less likely to develop and maintain protective immunity. While some studies suggested lower humoral responses in PWH [14], this did not appear to translate into a higher risk of re-infection in the pre-vaccine era, and several other studies did not find substantial differences following natural infection in PWH on suppressive ART [68–70]. Although some early data suggest that vaccine-induced immunity could be less robust among PWH, it appears to be driven by the population of PWH with low CD4+ cell counts [71].
Compared to antibody responses, data on cellular immune responses to SARS-CoV-2 infection in PWH are much more limited. A single high-quality study to date has shown similar T cell responses between PWH on suppressive ART and individuals without HIV [15]. In our rigorously matched natural infection cohort, we made three key observations regarding SARS-CoV-2-specific cellular immune responses in PWH compared to individuals without HIV. First, using peptide pools that include optimal SARS-CoV-2 epitopes that span the proteome, we found that PWH had lower SARS-CoV-2 specific CD8+ T cell responses. This difference was previously observed in non-Spike-specific T cell responses in PWH and may indicate that PWH have impaired capacity to mount a protective CD8+ T cell response upon SARS-CoV-2 re-infection, particularly with heterologous variants with immune-evading mutations in the Spike protein. It is also possible that PWH have expansion of other antigen-specific CD8+ T cells (e.g., CMV-specific) thereby diluting the SARS-CoV-2-specific pool as the denominator was total non-naive CD8+ T cells. Second, after controlling for multiple clinical factors including time of symptom onset, we found that SARS-CoV-2 specific CD4+ T cells had higher expression of the co-inhibitory receptor PD-1, suggesting they may have impaired functionality upon re-encountering infection. Alternatively, PWH may have more SARS-CoV-2 antigen exposure leading to a more exhausted cell phenotype. Third, we found that a higher CD4/CD8 ratio was associated with lower expression of PD-1 on SARS-CoV-2-specific CD8+ T cells among PWH. As higher CD4/CD8 ratios tend to be associated with ART initiation earlier in the course of infection [72], this finding adds to our understanding of how early HIV diagnosis and ART initiation is critical for preserving immune responses against heterologous infections in PWH.
This study has several limitations. The sample size was small, as relatively few PWH were infected with SARS-CoV-2 during the first year of the pandemic in our geographic area [73]. As there were few PWH without PASC, we were likely underpowered to make comparisons within the PWH sub-group; larger cohorts will be needed to do so. Our case definition, although potentially overly sensitive, is in line with consensus case definitions [16,17] and the association between HIV status and PASC was maintained in sensitivity analyses using more stringent definitions. However, given the nature of our recruitment, we emphasize that the high prevalence of PASC in our study cohort should not be considered to represent the population-level prevalence of this condition, which is likely much lower [27]. In addition, while the groups with and without HIV infection were well-matched on key variables demonstrated to be important in the immune response to SARS-CoV-2 infection and in PASC, the potential for residual confounding remains. For example, data on additional clinical factors that may be related to both HIV status and PASC, including comorbid mental health conditions and substance use, were unavailable. We used concurrent CD4+ T cell count and CD4/CD8 ratio as markers of immune reconstitution; CD4+ T cell nadir might be an important factor but we were unable to confirm these values in most participants due to fragmentation of healthcare records. Regarding laboratory measurements, we used a surrogate virus neutralization assay as opposed to a true neutralizing assay. Our T cell assay focused on IFNγ expression to identify SARS-CoV-2-specific T cells, which likely underestimates the magnitude of the total virus-specific response. We did not assess T cell proliferative capacity, which may reflect the capacity of virus-specific memory T cells to respond to a re-infection. Inflammatory marker data was not available on all participants, although sample availability for testing was based on the timing of the specimen collection and is not expected to bias the results. Finally, our population of PWH was mostly male, stable on ART, and had strong immune reconstitution, and these observations should be extended to populations with less access or adherence to ART or those with advanced HIV without further evidence in these subpopulations of PWH.
Although these findings need to be confirmed in larger and more diverse cohorts, our analysis provides compelling preliminary evidence that suggests an urgent need to better understand PASC within people living with HIV. Better understanding the epidemiology and pathophysiology of PASC within PWH may lead to targeted interventions to prevent or treat this newly identified condition among this special population of interest.
Data Availability
All data produced in the present study are available upon reasonable request to the authors.
FOOTNOTES
Author Contributions
MJP, JDK, JNM, SGD, and TJH designed the cohort, which was managed by RH and overseen by MJP. MJP, JDK, SL, RH, JNM, and SGD designed the study instruments. MJP, SEM, AFT, MIA, and RH recruited participants, coordinated research visits, administered study questionnaires, and collected clinical data. CAF, SM, AFT, SAG, and JYC performed data entry and quality control and addressed or adjudicated issues related to data integrity. SL cleaned the data, maintained the database, and oversaw data management. Antibody measurements were performed in the laboratory of KLL. NK, RLR, and TJH performed and/or oversaw the cellular immunology assays in the UCSF Core Immunology Laboratory. AC, JWW, and CJP performed and oversaw the inflammatory marker assays at Monogram Biosciences. DW and AS provided the SARS-CoV-2 peptide megapools. MJP, MAS, TMD, SEM, DVG, RLR, and TJH performed the analyses. MJP MAS, TMD, CAF, SEM, SM, SGD, RLR, and TJH drafted the manuscript with input from MSD, PYH, PWH, JFK, JNM, DVG, and MG providing in depth critical review. The study was funded by grants from MAS, MG, SGD, and TJH. All authors reviewed, edited, and approved the manuscript.
Funding
This work was supported by the National Institutes of Health (R01 AI141003, R01 AI158013, T32 AI60530, K23 AI157875, and K23 AI135037), and the UCSF/Gladstone Center for AIDS Research (CFAR). MJP received funding from the UCSF Resource Allocation Program and the CFAR Rapid COVID grant program. This work has been supported by NIH contract 75N93019C00065 (A.S, D.W).
Conflicts of Interest
AC, JWW, and CJP are employees of Monogram Biosciences. AS is a consultant for Gritstone Bio, Flow Pharma, Arcturus Therapeutics, ImmunoScape, CellCarta, Avalia, Moderna, Fortress and Repertoire. LJI has filed for patent protection for various aspects of T cell epitope and vaccine design work. TJH reports grants from Merck and Co. and Bristol-Myers Squibb outside the submitted work. The remaining authors have no conflicts related to the current work to report.
Acknowledgements
We are grateful to the study participants. We acknowledge current and former LIINC clinical study team members Tamara Abualhsan, Andrea Alvarez, Melissa Buitrago, Monika Deswal, Emily Fehrman, Heather Hartig, Yanel Hernandez, Marian Kerbleski, James Lombardo, Lynn Ngo, Enrique Martinez Ortiz, Antonio Rodriguez, Dylan Ryder, Ruth Diaz Sanchez, Viva Tai, Cassandra Thanh, Fatima Ticas, Leonel Torres, and Meghann Williams; and LIINC laboratory team members Amanda Buck, Joanna Donatelli, Jill Hakim, Nikita Iyer, Owen Janson, Christopher Nixon, Isaac Thomas, and Keirstinne Turcios. We thank the UCSF AIDS Specimen Bank for processing specimens and maintaining the LIINC biospecimen repository. The intracellular cytokine staining assays were performed in the UCSF Core Immunology Laboratory. We are grateful to Jon Oskarsson, Mary Shiels, Erin Pederson, Parousha Zand, and the UCSF Ward 86 and 360 Positive Health Practices for referring PWH who had COVID-19.
REFERENCES



