
Summary box
What is already known on this topic?COVID-19 mRNA vaccine efficacy wanes over time in vaccinated adults; a third vaccine booster is now recommended for individuals over 16 years of age six-months after the two-vaccine series is completed.
What is added by this report?We tested antibody durability of vaccine-induced full SARS-CoV-2 Spike and receptor binding domain (RBD) antibodies, as well as antibody levels specifically targeting the predominant variant of concern, Omicron, in younger adolescent children.
What are the implications for public health practice?Waning anti-SARS-CoV-2 antibodies in adolescents support urgent authorization for a COVID-19 mRNA booster in the 12-15-year-old age group.
Introduction
As we approach the third year of the COVID-19 pandemic, SARS-CoV-2 infection rates have surged due to highly infectious viral variants and waning population immunity. Vaccinated individuals are linked to increasing infections and hospitalizations, despite full (two-dose) mRNA vaccination(1). Boosters are recommended six months after full vaccination for individuals 16 years or older. The COVID-19 vaccine was approved for children ages 12-15 years old over six months ago, but boosters are not yet recommended for this age group.
While mortality from COVID-19 is lower in children compared to adults, over 7.5 million children have been diagnosed with COVID-19, leading to 28,000 hospitalizations(1) and 12,000 pediatric deaths worldwide(2). Additionally, 6,000 children in the US have been diagnosed with the post-infectious illness, Multisystem inflammatory syndrome in Children (MIS-C)(3), and more than one in seven adolescents experience long COVID-19(4). Children are not spared from infection and vaccination remains a critical strategy for preventing infection, transmission, severity of disease, and complications of COVID-19. Thus, we must understand and optimize vaccine responses across all pediatric age groups. Here, we quantified antibody responses immediately following the Pfizer-BioNTech mRNA vaccination and six months post-inoculation against the wildtype SARS-CoV-2 Spike, RBD and new Omicron Variant of Concern (VOC).
Methods
Adolescent children and their parents assented and consented, respectively, to participate in the MGH Pediatric COVID-19 Biorepository (MGB IRB #2020P000955)(5). Demographic information was extracted from medical records. Blood was collected into serum separation tubes by venipuncture or by microneedle device, prior to vaccination (V0), 2-3 weeks after the initial Pfizer-BioNTech mRNA vaccination (V1), 2-4 weeks after the second dose (V2), and again 6 months after the vaccination series was complete (V6), with a subgroup only having the V6 collection. Blood was allowed to clot then spun by centrifuge. Serum was collected and serological analyses were performed using an in-house enzyme-linked immunosorbent assay (ELISA) that detects IgG against the wildtype SARS-CoV-2 Spike, the wildtype RBD, or the Omicron SARS-CoV-2 VOC RBD by using the previously described method(6). Briefly, 384-well plates were coated with 0.5µg/ml of Spike or RBD overnight at 4°C in bi-carbonate buffer. The plates were then washed and plasma samples were added at a 1:500 (Spike) dilution or 1:200 (RBD) dilution in duplicate for 1h at 37°C, washed and then detected with a secondary anti-IgG (Bethyl Laboratories). The secondary was washed away after 1h, and the colorimetric detector was added (TMB; Thermo Fisher) for 5 mins, the reaction was stopped and the absorbance was acquired at 450/570nm on SpectraMax plate reader. In order to convert raw OD values into concentration, a two-fold dilution curve (starting at 29.8 international units) of the WHO standard (NIBSC code: 20/136) was included onto every ELISA plate. The sample concentration was interpolated from the resulting standard curve, as previously described(6). Analysis was completed by Prism 9.3 using one-way ANOVA for multiple comparisons and t-test for two-way comparisons.
Results
Thirty-one children enrolled in our study, nineteen of whom provided blood samples at four separate time points: prior to vaccination (V0), 2-3 weeks after first vaccine dose (V1), 2-4 weeks after the second vaccine dose (V2), and 6 months after the second vaccine dose (V6). The median age of the cohort was 13.9 years; 27 children were between the ages of 12-15 years; four were between ages of 16-19 years. Sex was equally distributed, and one quarter of the population was Hispanic (Table 1). All participants denied SARS-CoV-2 infection prior to enrollment and throughout the course of the study.
Demographics of adolescent participants and timing of sample collections.
Antibody responses to wildtype SARS-CoV-2 Spike (Figure 1A), wildtype SARS-CoV-2 RBD (Figure 1B), and SARS-CoV-2 Omicron variant RBD (Figure 1C) were analyzed at each time point. Robust generation of anti-SARS-CoV-2 antibodies was seen following the second dose of the vaccine (P > 0.0001), followed by a significant loss of antibody response after six months. While a few adolescents maintained high levels of anti-Spike or anti-RBD antibodies at six months (V6), most children exhibited similar antibody levels to those detected after the first vaccine dose (V1), or in the case of anti-Spike antibodies, pre-vaccine levels of antibodies (V0). This loss of antibody levels highlights a potential vulnerability of adolescents to breakthrough infections six months after their vaccine series.

Humoral responses to a) Wildtype (WT) Spike b) WT Receptor Binding Domain (RBD), and c) Omicron RBD are quantified prior to vaccination, 2-3 weeks following the first vaccine dose, 2-4 weeks following the second mRNA vaccine dose, and 6 months following the second mRNA vaccine dose. Displayed as international units. Analysis by ANOVA. ns = not significant, * P < 0.05, *** P < 0.001, **** P < 0.0001
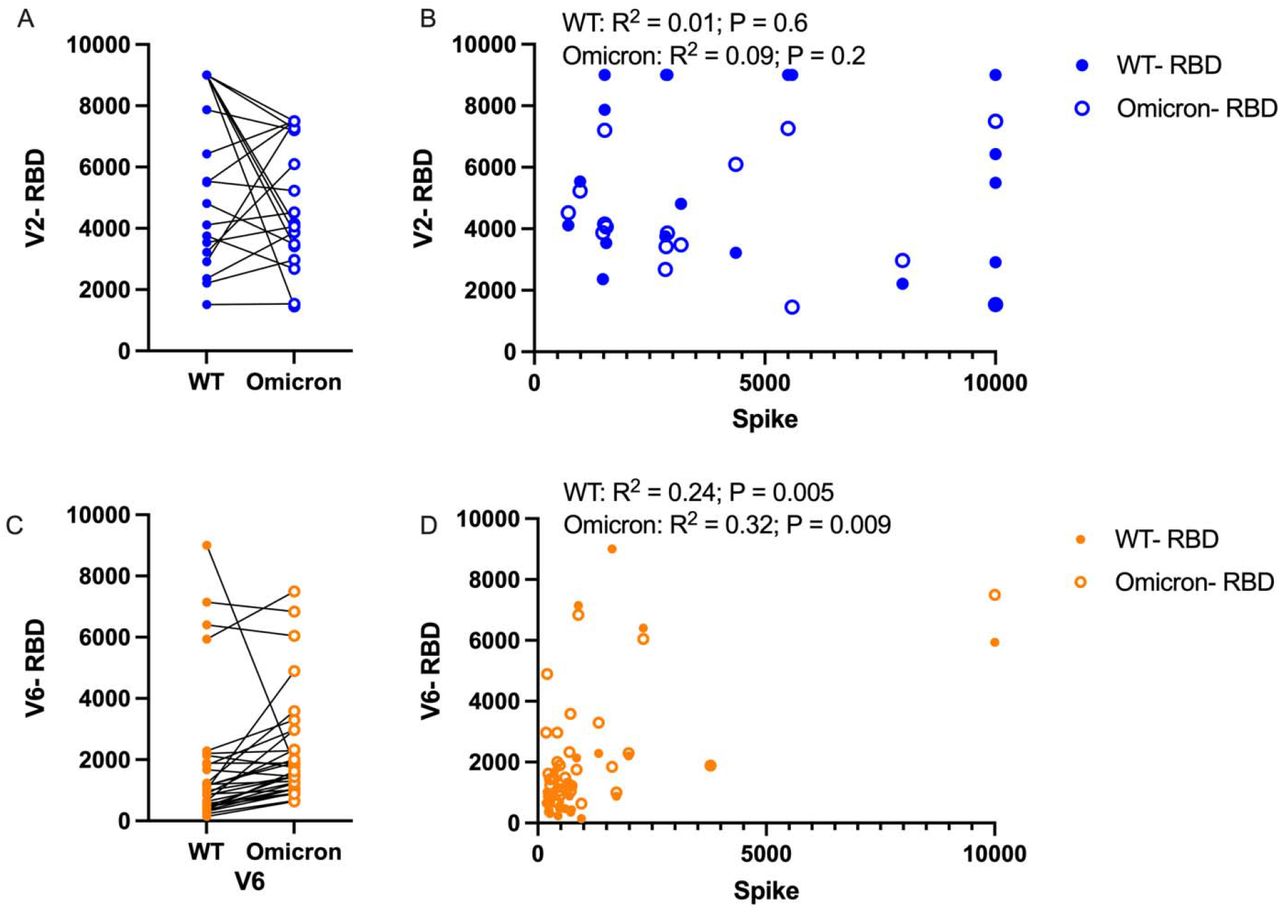
As Omicron is likely to become the predominant SARS-CoV-2 variant globally, we assessed whether adolescent children displayed coverage and protection against the Omicron-specific SARS-CoV-2 RBD. Interestingly, while a subgroup of adolescents showed a drop in sensitivity towards Omicron as compared to wildtype in the weeks following the second vaccine, most adolescents displayed equal sensitivity for Omicron and wildtype RBD after the second vaccine (Figure 2A). At peak immunity following V2, there was no correlation in Spike or RBD for both wildtype and Omicron (Figure 2B). At six-months, adolescents even displayed a trend towards increased immune responses against Omicron, despite the overall trend to lower total antibody titers (Figure 2C). Lastly, a strong correlation was seen in declining anti-Spike and anti-RBD titers for both wild type and Omicron (Figure 2D).
{kind=link}
{kind=link}
A) Following the second mRNA vaccine dose, anti-RBD responses titers are compared between WT and Omicron, B) and correlations between RBD and Spike were assessed for each variant. C) Anti-RBD titers were also compared at the 6-month time point, and correlation between RBD and Spike was again assessed. Paired analysis with t-test, correlation with Pearson correlation.
Discussion
As the COVID-19 mRNA vaccines represent a new vaccination platform, the longevity of immune responses needs to be characterized across all age ranges, especially in light of emerging variants. Here, we specifically focus on humoral immune responses in adolescent children against SARS-CoV-2, including responses against the highly infectious and predominant variant of today, Omicron.
As expected and as seen in adult populations, mRNA vaccine-induced immunity wanes over a 6-month time period in adolescent children to near pre-vaccination levels. This finding demonstrates a vulnerability for infection in adolescent children ages 12-15 years, many of whom have now received their vaccine series over six months ago. Encouragingly, the immune responses that have been generated display sufficient cross-coverage of the VOC, Omicron, with stronger immune responses than have been reported in adults(7). It is possible that Omicron responses are overestimated in our assay as the standards are normalized for wildtype. Alternatively, this could suggest that children develop a more adaptive humoral immune response than adults(8), which may benefit children as additional variants of concern emerge. While it is plausible that vaccine-induced anamnestic immunity may provide some level of protection in this population upon exposure, the durability of vaccine-induced immunity wanes in adolescents, and boosting may promote a robust barrier of immunity that will contribute to public health efforts to limit spread and prevent future hospitalizations and severe disease.
In conclusion, adolescent children exhibit waning antibody immune responses six months post-mRNA vaccine. Authorization for mRNA boosters is urgently needed in the 12-15-year-old age group to bolster durable immune responses in the face of the surging COVID-19 pandemic.
Data Availability
All data produced in the present study are available upon reasonable request to the authors
Acknowledgements
We thank the children and families who participated in this research. We thank from Massachusetts General Hospital for Children, Nancy Zimmerman, Mark and Lisa Schwartz, an anonymous donor (financial support), Terry and Susan Ragon, and the SAMANA Kay MGH Research Scholars award for their support. We acknowledge support from the Ragon Institute of MGH, MIT, and Harvard, the Massachusetts Consortium on Pathogen Readiness (MassCPR), the NIH (3R37AI080289-11S1, R01AI146785, U19AI42790-01, U19AI135995-02, U19AI42790-01, 1U01CA260476 – 01, CIVIC75N93019C00052, 5K08HL143183, R01HD100022-02S2), the Gates Foundation Global Health Vaccine Accelerator Platform funding (OPP1146996 and INV-001650), and the Musk Foundation.



