
Abstract
Vaccines based on the spike protein of SARS-CoV-2 are a cornerstone of the public health response to COVID-19. The emergence of hypermutated, increasingly transmissible variants of concern (VOCs) threaten this strategy. Omicron, the fifth VOC to be described, harbours 30 amino acid mutations in spike including 15 in the receptor-binding domain. Here, we demonstrate substantial evasion of neutralisation by Omicron in vitro using sera from vaccinated individuals. Importantly, these data are mirrored by a substantial reduction in real-world vaccine effectiveness that is partially restored by booster vaccination. We also demonstrate that Omicron does not induce cell syncytia and favours a TMPRSS2-independent endosomal entry pathway. Such marked changes in antigenicity and replicative biology may underlie the rapid global spread and altered pathogenicity of the Omicron variant.
Main
Vaccination against SARS-CoV-2 is based primarily on vaccines that induce immunity to the spike glycoprotein. These vaccines have become the cornerstone of the global public health response to SARS-CoV-21. However, their effectiveness is now being threatened by the emergence of Variants of Concern (VOC) displaying enhanced transmissibility and evasion of host immunity2. Of the five VOCs that have emerged, the Beta variant (B.1.351) and Gamma (P.1) variants were associated primarily with immune evasion; they spread locally but never dominated globally. In contrast, the Alpha (B.1.1.7) and Delta (B.1.617.2) VOCs spread globally and were responsible for significant waves of infections and an increase in reproduction number (R0). The Alpha and Delta variants harbour mutations within the polybasic cleavage site in spike (a H681 in Alpha and R681 in Delta) that enhance cleavage by furin; changes that are associated with enhanced cell entry and may contribute to increased transmissibility. While the Alpha variant spread rapidly, it was in turn replaced by the Delta variant that combined augmented transmissibility with significant immune evasion2–5.
Omicron (lineage B.1.1.529) is the fifth variant to be named as a VOC by the World Health Organisation (WHO) and was first detected in mid-November 2021 in Botswana, South Africa6 and in quarantined travellers in Hong Kong7. It has since split into three divergent sublineages (BA.1, BA.2 and BA.3) of which BA.1 now dominates worldwide.
Emerging data indicate that the Omicron variant evades neutralisation by sera obtained from people vaccinated with 1 or 2 doses of vaccine, especially when antibody titres are waning. Indicative studies have shown that 3 doses of spike-based vaccines may provide only partial protection from infection with this variant, including unpublished data made available as a press release from Pfizer. Immune evasion by Omicron may have contributed to the extremely high transmission rates in countries with high vaccination rates or natural immunity (R0 of 3-5 in the U K 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18.
In this study, we investigate the antigenic and biological properties of the Omicron variant that might underlie immune evasion and increased transmission of the virus using both in vitro assays and real-life population data.
Results
Omicron displays substantial changes within spike predicted to affect antigenicity and furin cleavage
Omicron is characterised by significant changes within the RBD of the spike glycoprotein, regions targeted by class 1,2 and 3 RBD-directed antibodies, and within the N-terminal domain (NTD) supersite (Fig.1a). The G339D, N440K, S477N, T478K, Q498R and N501Y mutations enhance binding of spike to the human ACE2 receptor, while combinations such as Q498R and N501Y may enhance ACE2 binding additively19. Overall, the Omicron RBD binds to the human ACE2 with approximately double the affinity (x2.4) of the Wuhan RBD8. Deep mutational scanning (DMS) estimates at mutated sites are predictive of substantially reduced monoclonal and polyclonal antibody binding and altered binding to human ACE2 (Fig.1b)20. Fourteen mutations (K417N, G446S, E484A, Q493R, G496S, Q498R and to a lesser extent, G339D, S371L, S373P, N440K, S477N, T478K, N501Y and Y505H) may affect antibody binding based on a calculated escape fraction (a quantitative measure of the extent to which a mutation reduces polyclonal antibody binding by DMS). Seven Omicron RBD mutations (K417N, G446S, E484A, Q493R, G496S, Q498R and N501Y) have been shown previously to be associated with decreased antibody binding, importantly falling in epitopes corresponding to three major classes of RBD-specific neutralising antibodies (nAbs). The mutations present in spike also involve key structural epitopes targeted by several monoclonal antibodies in current clinical use. Of these, bamlanivimab, cilgavimab, casirivimab, etesevimab, imdevimab, regdanvimab and tixagevimab bind to the RBM, and neutralisation of Omicron has been shown to be negligible or absent. In contrast, sotrovimab, targets a conserved epitope common to SARS-CoV-1 and SARS-CoV-2 that is outside the RBM and has only a small reduction (x3) in neutralisation potency21–23. N679K and P681H mutations at the furin cleavage site (FCS) are predicted individually to increase furin cleavage, although the combination of these changes and an adjacent change (H655Y, also present in the Gamma VOC) in the vicinity of the FCS is unknown24.
Omicron bears three deletions (amino acids 69-70, 143-145 and 211) and an insertion (site 214) in the NTD of spike. The 69-70 deletion is also found in the Alpha and Eta (B.1.525) variants and is associated with enhanced fusogenicity and incorporation of cleaved spike into virions25. This 69-70 deletion is a useful proxy for estimates of Omicron prevalence in the population by S-gene target failure (SGTF) using the TaqPathTM (Applied Biosystems, Pleasanton, CA) diagnostic assay. Deletions in the vicinity of amino acids 143-145 have been shown to affect a range of NTD-specific nAbs26, 27.
Emergence of the Omicron variant in the UK
Despite high vaccination rates and levels of natural immunity following previous exposure in the UK, Omicron has rapidly become dominant. The evolutionary relationships of SARS-CoV-2 variants at a global level are shown in Fig.1c. The first 8 cases of Omicron were detected in the UK on the 27th and 28th November 2021 (2 in England and 6 in Scotland). Due to the rapid spread of Omicron, early genome sequences were highly related with an average genetic divergence between 1 and 7 single nucleotide polymorphisms (SNPs) (Fig.1d). The phylogenetic relationship to Omicron sequences from other countries was consistent with multiple introductions associated with travel to South Africa followed by community transmission.

a, Spike homotrimer in open conformation with locations of Omicron amino acid substitutions, deletions (Δ), or insertions (ins) highlighted as spheres with opaque surface representation. Colouring highlights mutations at residues with substitutions impacting RBD-specific antibodies of classes 1 (green), 2 (yellow), and 3 (blue)28, or that belong to the NTD antibody supersite (magenta)26, or that belong to the FCS (orange), with the remainder in grey. These are annotated on the monomer with an ‘up’ receptor-binding domain. The substitution D614G which is shared by common descent by all lineage B.1 descendants is italicised. The visualisation is made using a complete spike model29 which is in turn based upon a partial cryo-EM structure (RCSB Protein Data Bank (PDB) ID: 6VSB30. b, Aligned heatmaps showing properties of amino acid residues or of the specific amino acid substitution present in the Omicron variant, as appropriate (insertion not shown). Structure-based epitope scores31 for residues in the structure of the original genotype spike in closed and open conformations are shown. For RBD residues, the results of deep mutational scanning (DMS) studies show the escape fraction (that is, a quantitative measure of the extent to which a mutation reduced polyclonal antibody binding) for each mutant averaged across plasma (‘plasma average’) and for the most sensitive plasma (‘plasma max’)20. Each mutation is classified as having evidence for mutations affecting neutralisation by either mAbs27, 32–35 or antibodies in convalescent plasma from previously infected or vaccinated individuals20, 34–36. Membership of the furin cleavage site is shown. The distance to ACE2-contacting residues that form the receptor-binding site (RBS) is shown (RBS defined as residues with an atom <4Å of an ACE2 atom in the structure of RBD bound to ACE2 (RCSB PDB ID: 6M0J37. Finally, ACE2 binding scores representing the binding constant (Δlog10 KD) relative to the wild-type reference amino acid from DMS experiments38. c, Inferred evolutionary relationships of SARS-CoV-2 from NextStrain (https://nextstrain.org/ncov/gisaid/global) with the Variants of Concern labelled. The colours of the tree tips correspond to the number of mutations causing Spike amino acid substitutions relative to the SARS-CoV-2 original genotype (OG) reference strain Wuhan-Hu-1. d, Inferred evolutionary relationships of the first 111 Omicron sequences in Scotland with NHS Scottish Health boards denoted: AA, Ayrshire and Arran; FF, Fife; FV, Forth Valley; GC, Great Glasgow and Clyde; GR, Grampian; HG, Highlands; LN, Lanarkshire; LO, Lothian; TY, Tayside, see key.
Neutralising responses to Omicron (BA.1) are substantially reduced following double and partially restored following triple vaccination
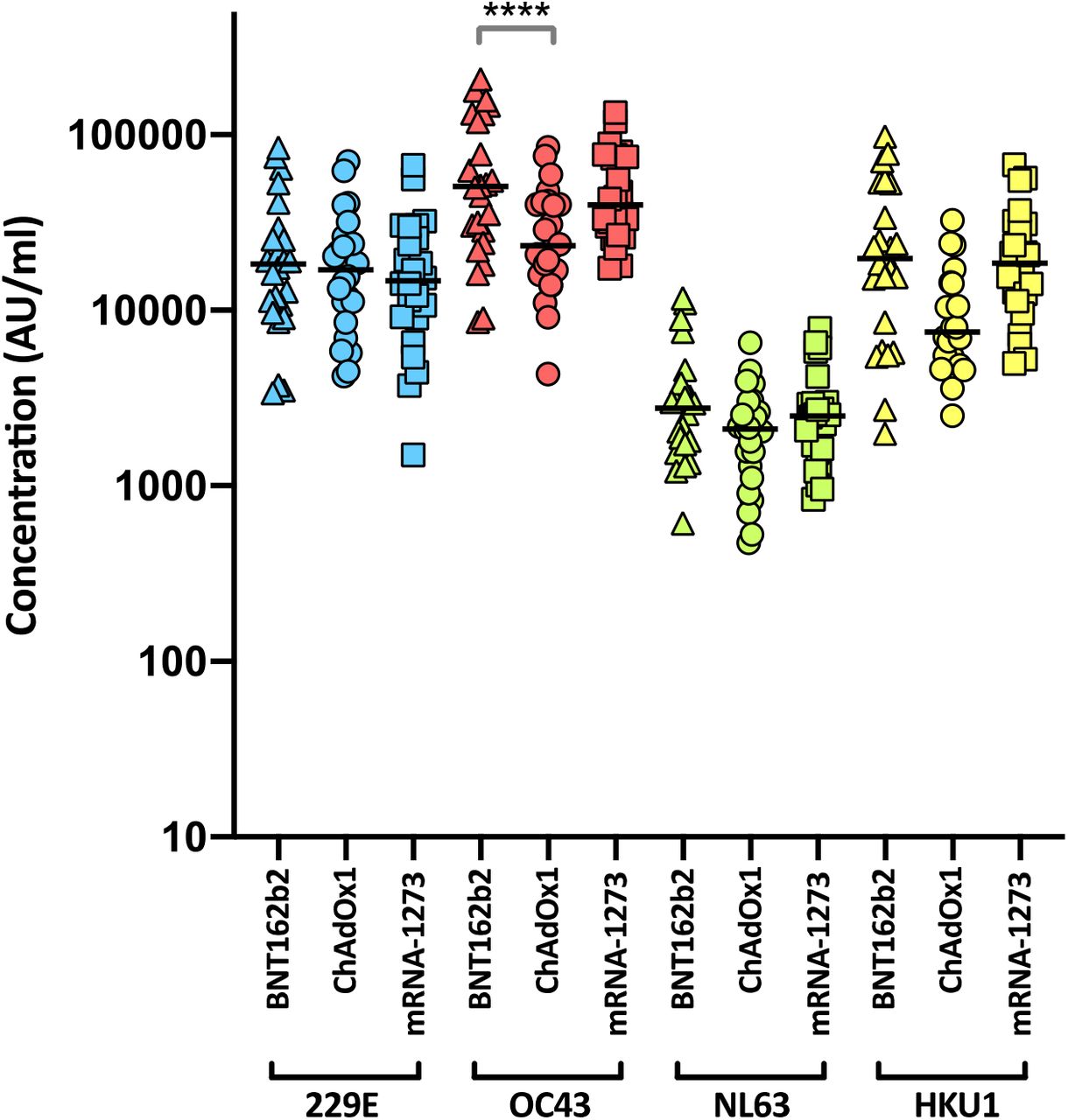
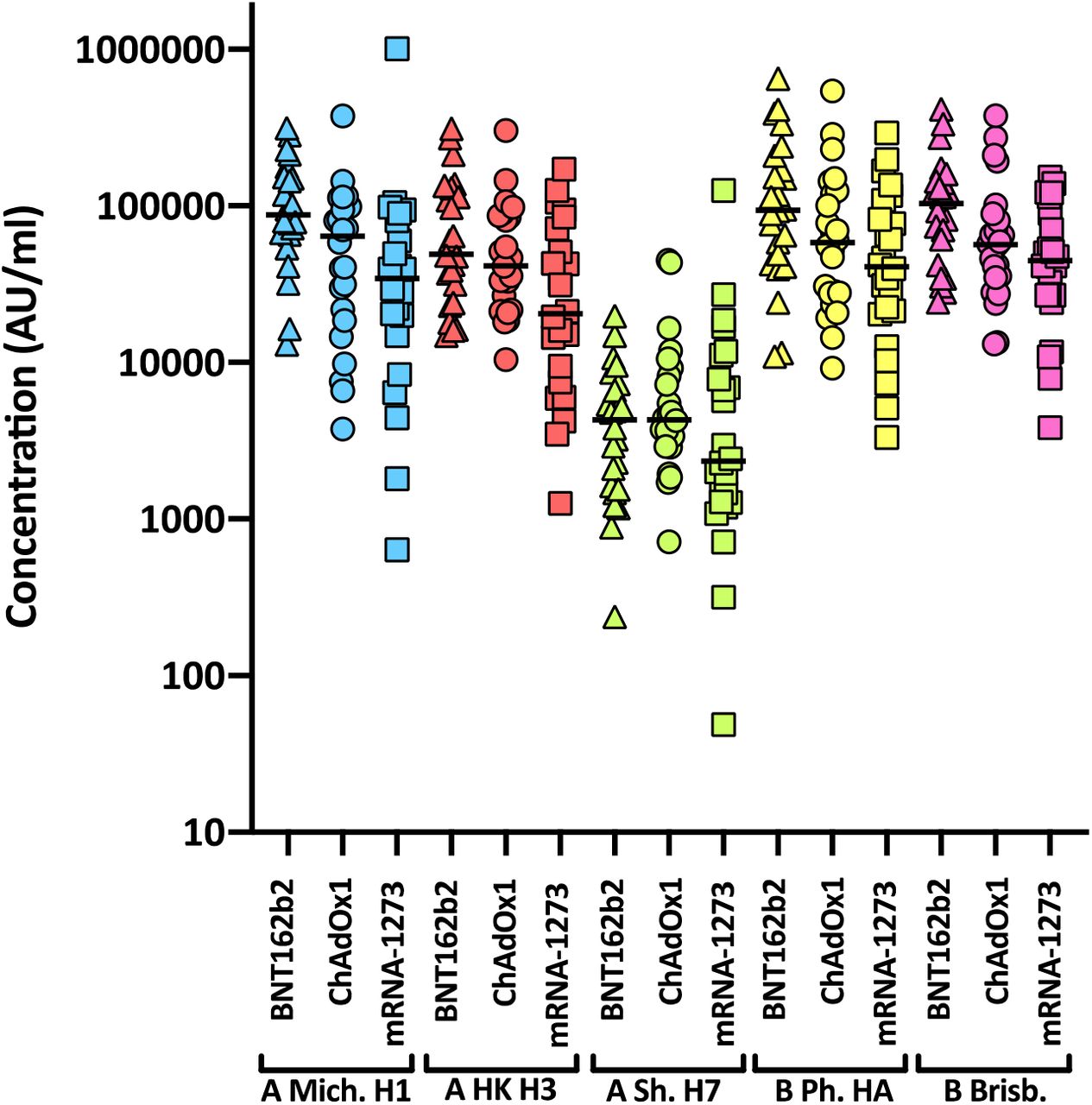
Levels of nAbs in patient sera correlate strongly with protection from infection39–42, and reductions in neutralising activity against the Alpha and Delta variants are consistent with a reduction in vaccine effectiveness2–5, 43. To predict the effect of the mutations within the Omicron spike glycoprotein on vaccine effectiveness, sera collected from healthy volunteers at more than 14 days post-2nd dose vaccination with either BNT162b2, ChAdOx1 or mRNA-1273 were sorted into three age-matched groups (n=24 per group, mean age 45 years). Sera were first screened by electrochemiluminescence (MSD-ECL) assay for reactivity with SARS-CoV-2 antigens (Spike, RBD, NTD or nucleoprotein (N)). The antibody responses to RBD and NTD were significantly higher (p<0.0001) in the sera from individuals vaccinated with BNT162b2 or mRNA-1273 in comparison with the ChAdOx1 vaccinees (Fig. 2a and Supplementary Table 1). In contrast, antibody responses to endemic human coronaviruses (HCoVs) (Extended data Fig. 1 and Supplementary Table 2) or influenza (Extended data Fig. 2 and Supplementary Table S3) were similar, with the exception of coronavirus OC43, where responses in BNT162b2 and ChAdOx1 vaccinees differed significantly, perhaps suggesting modulation (back-boosting) of pre-existing OC43 responses by BNT162b2 vaccination.

Antibody responses were studied in three groups of individuals (n=24 per group) receiving primary vaccination with either BNT162b2, ChAdOx1 or mRNA-1273 by a, MSD-ECL assay or b, pseudotype-based neutralisation assay. a, Responses were measured against full-length spike glycoprotein (Spike), receptor binding domain (RBD), N-terminal domain (NTD) and nucleoprotein (N) and are expressed as arbitrary units (AU/ml). b, NAb responses were quantified against Wuhan or Omicron spike glycoprotein bearing HIV (SARS-CoV-2) pseudotypes. Each point represents the mean of three replicates, bar represents the group mean. To assess the effect of booster vaccines antibody responses were studied in two groups of individuals primed with two doses of either BNT162b2 or ChAdOx1 and boosted with either BNT162b2 or mRNA-1273. Reactivity against SARS-CoV-2 antigens was measured by c, MSD-ECL assay while neutralising activity d, & e, was measured using HIV (SARS-CoV-2) pseudotypes, as above. Green data points represent those boosted with mRNA-1273, all others received BNT162b2. In panel d, & e, % Omicron neutralising refers to the proportion of serum samples that displayed neutralising activity against Omicron pseudotypes.
Next, the nAb responses against SARS-CoV-2 pseudotypes expressing the spike glycoprotein from either Wuhan-Hu-1, or Omicron (BA.1) were compared (Fig. 2b). Vaccination with mRNA-1273 elicited the highest nAb titres (mean titre Wuhan=21,118, Omicron=285), in comparison with those elicited by vaccination with either BNT162b2 (Wuhan=4978, Omicron=148.3) or ChAdOx1 (Wuhan=882.3, Omicron=61.9). Neutralising antibody titres against Wuhan differed significantly between the three study groups. Activity against Omicron was markedly reduced in comparison with Wuhan, reduced by 33-fold for BNT162b2, 14-fold for ChAdOX1 and 74-fold for mRNA-1273 (Supplementary Table 4). While the fold change in neutralisation was lowest in recipients of the ChAdOx1 vaccine and highest in recipients of the mRNA-1273 vaccine, absolute neutralisation values were highest in mRNA-1273 followed by BNT162b2 and ChAdOx1. Neutralisation was lowest in the ChAdOx1 group, however it is important to note that this was given to older patients during early vaccine rollout in the UK, especially to vulnerable patients in nursing homes and was not recommended in young adults less than 40 years of age.
Next, samples were analysed from vaccine recipients at least 14 days post booster vaccination (third dose). Participants had been primed with two doses of either ChAdOx1 or BNT162b2, followed by a third dose of either BNT162b2 (full dose) or mRNA-1273 (half dose; 50µg). All sera reacted strongly with SARS-CoV-2 antigens by MSD-ECL, with no significant differences between the four groups (Fig. 2c and Supplementary Table 5). Antibody responses to HCoVs (Extended data Fig. 3 and Supplementary Table 6) or influenza (Extended data Fig. 4 and Supplementary Table 7) were similar, with the exception of influenza Michigan H1, where responses in ChAdOx1-primed and BNT162b2 or mRNA-1273-boosted groups differed significantly, likely reflecting co-administration of influenza booster vaccines during the booster campaign. Two vaccine recipients boosted with BNT162b2 displayed weak reactivity with nucleocapsid (Fig. 2c), suggesting previously undetected exposure to SARS-CoV-2. Sera from vaccine recipients primed with BNT162b2 and boosted with either BNT162b2 or mRNA-1273 displayed similar titres of nAb against Wuhan to the samples collected post-dose 2 (Fig. 2d). In contrast, vaccination of individuals primed with ChAdOx1 with a booster dose of either BNT162b2 or mRNA-1273 resulted in a marked increase in antibody titre (9.3-fold increase) against Wuhan relative to the low titres after dose 2 (Fig. 2e and Supplementary Table 8). The marked increase in antibody titre in ChAdOx1-primed individuals (Extended data Fig. 5) emphasises the importance of the third dose booster in this population. Indeed, following boost with either BNT162b2 or mRNA-1273, anti-Wuhan nAb titres in the ChAdOx1-primed group were not significantly different from those primed with BNT162b2 (Supplementary Table 8). NAb titres against Omicron were lower in both booster study groups and did not differ significantly in titre (Supplementary Table 8). However, absolute numbers displaying measurable Omicron neutralising activity were higher in the ChAdOx1-primed group (13/21, 62%) compared with the BNT162p2 primed group (5/20, 25%) (Fig. 2d, Fig. 2e).
Vaccine effectiveness against the Omicron variant is reduced compared to the Delta variant
A logistic additive model with a test negative case control design was used to estimate relative vaccine effectiveness against becoming a confirmed case with Delta (4911 cases) and/or Omicron (6166 cases) in a population of 1.2 million people in the largest health board in Scotland, NHS GG&C, between 22nd and 28th December 2021. Demographic data are shown in Supplementary Table 9. The timing of first doses of vaccination are shown in Fig.3a and the occurrence of sequenced/confirmed infections with different variants in vaccine recipients over time is shown in Fig.3b. Infection status for Omicron and Delta was modelled by number and product type of vaccine doses, previous infection status, sex, SIMD quartile, and age (to control for demographic bias). Immunosuppressed individuals were removed from the analysis to ensure case-positivity could be attributed to vaccine escape rather than an inability to mount a vaccine response, with immunosuppression status derived from a list of those in GG&C shielding due to immunosuppression or listed as being given immunosuppressant drugs. Age and time since vaccination were each modelled as single smooth effects using thin plate regression splines44.

a, Denominator (violin) plot showing populations of test positive and test negative cohorts in NHS GG&C, with the widths of the grey bands represent the populations in each group at each time point. VOC classification of sequenced cases are overlaid as a dotplot, with points coloured by their VOC and a random jitter applied along the x-axis for visual clarity. b, Boxplots of date of first administered vaccine dose by vaccine product for the population of NHS Greater Glasgow and Clyde (NHS GG&C) aged 18 and older. The box limits are the quartiles and the centre line is the median, with whisker length of 1.5 times the interquartile range. Outliers are shown as dots outwith the whisker range. Data points are overlaid as a dotplot with points shown as black dots, with a random jitter along the x-axis applied for visual clarity. c, Error bar plot of estimated vaccine effectiveness against testing positive for Delta and Omicron SARS-CoV-2 infection in the population of over 18s in NHS GG&C who were tested between 6th and 12th December 2021. The points and corresponding text represent the estimated vaccine effectiveness (%) for each group, for each variant, with the error bar endpoints representing the endpoints of the corresponding 95% CIs. The additive effect of infection-acquired immunity was calculated for the entire population and plotted for the unvaccinated cohort.
The protection from vaccine-acquired and infection-acquired immunity were estimated as being markedly reduced against Omicron compared with Delta. Estimates of vaccine effectiveness in recent recipients (at 14 days post-dose) were only 11.44% for full primary courses of ChAdOx1 against Omicron and 78.92% against Delta. For two doses of mRNA vaccines, vaccine effectiveness was significantly lower for Omicron than Delta; BNT162b2 (26.02% versus 83.55%) and mRNA-1273 (23.70% versus 87.82%) (Fig.3c). These responses were similar following a third booster dose of BNT162b2 or mRNA-1273 against Delta (85.94% and 86.53%, respectively), but increased significantly against Omicron (43.22% and 46.38%, respectively). These estimates are in keeping with those reported recently against symptomatic infection in England where vaccine effectiveness was estimated as 71.4% and 75.5% for ChAdOx1 and BNT162b2 primary course recipients boosted with BNT162b2, respectively9.
Next, we estimated the additive protective effect of previous natural infection. Infection-acquired immunity directed against other VOCs may be broader in nature and may wane more slowly than that induced by vaccines45–47. The level of protection following previous infection was 33.9% for Omicron, and 77.4% for Delta. This level of protection was greater than that observed following two doses of vaccine for Omicron but did not reach the levels attained by those who had never had natural infection and had received third dose boosters for either Omicron or Delta. Collectively, these results emphasise the importance of booster vaccines. Importantly, vaccine-mediated protection against severe disease is likely to be more durable than that against detected infection48.
Absence of syncytia in Omicron-infected cells
Our data demonstrate that antigenic change in Omicron permits evasion of vaccine induced immunity, however, the constellation of spike mutations in Omicron suggest that functional change may also contribute to its rapid transmission (Fig.1a). Therefore, we investigated the virological properties of live Omicron isolated from a patient sample. SARS-CoV-2 particles can achieve membrane fusion at the cell surface following proteolytic activation of spike by the plasma membrane protease TMPRSS2. This property also permits spike-mediated fusion of SARS-CoV-2 infected cells with adjacent cells resulting in syncytia49; a feature that has been associated with severe disease50. Moreover, the Delta variant has been shown to exhibit enhanced fusion compared to the Alpha and Beta variants51.
A split GFP cell-cell fusion system52 was used to quantify syncytia formation n by Omicron, Delta and first wave Wuhan D614G virus (Fig. 4a). Cells expressing split GFP were infected with Wuhan-D614G, Delta or Omicron and the levels of the reconstituted GFP signal following cell-cell fusion was determined in real time (Fig. 4c). In addition, infected cells were probed by indirect immunofluorescence assay to assess viral replication by the detection of the viral nucleocapsid protein (Fig. 4b). The Delta variant exhibited the highest levels of cell fusion followed by Wuhan D614G. In contrast, the Omicron variant failed to form syncytia. This failure was not due to lack of infection as immunofluorescent detection of nucleocapsid protein confirmed viral replication by Omicron, Wuhan D614G and Delta18.

a, Schematic representation of the split GFP system, used in this study to quantify virus induced cell fusion. This system is based on co-culture of two different cell lines (GFP-10 and GFP-11) expressing split GFP molecules. Upon virus-induced cell fusion, the intact GFP molecule is reconstituted, and the resulting signal can be detected and quantified. b, GFP-10 and GFP-11 A549 ACE2 TMPRSS2 cells were co-cultured and infected with Wuhan D614G, Delta and Omicron and incubated in a CLARIOStar Plus (BMG LABTECH) at 37°C / 5% CO2. Micrographs display cells 22 hours post infection: reconstituted GFP (green), N viral nucleocapsid (red), and DAPI nuclear counter stain (blue). Scale bars 100µm. c, To quantify, GFP signal was measured every 30 min for 20h. Omicron infected cells showed only background levels of GFP signal. d, Calu-3 cells were infected with Wuhan D614G, Delta and Omicron and supernatants were collected at the indicated times and assessed by RT-qPCR. Omicron display reduced replication kinetics compared to Wuhan D614G and Omicron. e, SARS CoV-2 entry can occur via two routes. Route 1 permits rapid fusion at the cell surface following proteolytic processing by TMPRSS2. In Route 2 fusion occurs following endocytosis after processing by cathepsin B or L. Route 1 and 2 can be specifically inhibited using the protease inhibitors Camostat and E64d, respectively. f, SARS-CoV-2 pseudotype infection of the stated cell lines, data represent mean luciferase values from one representative experiment. In Calu-3 cells Route 1 entry predominates whereas HEK exclusively support Route 2, A549 ACE2 TMPRSS2 cells permit both routes. Pangolin CoV spike is included as a control; it can only achieve entry via Route 2. Pseudotypes without viral glycoproteins (No) are included as a negative control. g, Relative SARS-CoV-2 pseudotype infection (compared to untreated control) of cells treated with 10μM protease inhibitors. Data represent mean of four replicates, error bars indicate standard error of the mean, asterisks indicate statistical significance (ANOVA). h, Titration of Camostat and E64d against Delta, Omicron and Pangolin CoV in A549 ACE2 TMPRSS2 cells, data points represent mean relative infection, compared to untreated control.
Reduced replication kinetics of Omicron in lung epithelial cells
The replication of Omicron, Delta and Wuhan D614G was compared in Calu-3, a human lung epithelial cell line. Wuhan D614G and Delta displayed comparable replication kinetics over a period of 72 hours, with visible CPE between 48-72hpi (Fig. 4d). In contrast, the titres of Omicron were at least an order of magnitude lower at each time point compared to Wuhan D614G and Delta. These observations are consistent with attenuated replication of Omicron in lower respiratory tissues as recently reported18, 53.
Omicron spike has switched entry route preference
Entry of SARS-CoV-2, and related coronaviruses, can proceed via two routes54. Cell surface fusion following proteolysis by TMPRSS2, as described above (“Route 1”, Fig. 4e), or fusion from the endosome after endocytosis and activation by the endosomal proteases Cathepsin B or L (“Route 2”, Fig. 4e). The ability of SARS-CoV-2 to achieve cell surface fusion is dependent on its S1/S2 polybasic cleavage site; this is absent from most closely related sarbecoviruses, which are confined to endosomal fusion55–57. Given the reduced fusogenicity and replication kinetics of Omicron, HIV pseudotypes were used to evaluate entry route preference. We tested Wuhan D614G, Alpha, Delta and Omicron spike, while Pangolin CoV (Guangdong isolate) spike was included as a control. Pangolin CoV spike exhibits high affinity interactions with human ACE2 but lacks a polybasic cleavage site and, therefore, enters via the endosome only58–61.
Calu-3 cells support cell surface (Route 1) fusion predominantly, owing to their high endogenous expression of TMPRSS256, 62; in these cells, Delta yielded the highest infection, being ∼4 fold higher than Omicron (Fig. 4f). Pangolin CoV infection was low, indicating that Calu-3 cells do not support robust endosomal entry. In contrast, HEK only support endosomal entry and in these cells Pangolin CoV had high infection. Notably, Omicron also achieved high infection in HEK cells, producing ∼10-fold greater signal than Delta. This suggests that Omicron, like Pangolin CoV, is optimised for endosomal entry. All pseudotypes exhibited robust infection in A549-ACE2-TMPRSS2, where both entry routes are available63, 64.
Entry pathway preference was further investigated using protease inhibitors targeting either TMPRSS2 (Camostat) or cathepsins (E64d)55. In Calu-3 cells, all SARS-CoV-2 pseudotypes were inhibited by Camostat, whereas only Omicron exhibited E64d sensitivity, indicating that a component of infection occurs via endosomal entry (Fig. 4g). In HEK cells, all pseudotypes were inhibited by E64d, whereas Camostat was non-inhibitory; this confirms that only endosomal entry is available in these cells. Inhibitor treatment in A549 ACE2 TMPRSS2 provided the clearest evidence of altered entry by Omicron. D614G, Alpha and Delta were potently inhibited by Camostat, but not E64d. For Omicron, and Pangolin CoV, this pattern was reversed, suggesting a binary switch from cell surface to endosomal fusion; this conclusion was supported by titration of either inhibitor in A549 ACE2 TMPRSS2 cells (Fig. 4h).
These data indicate that, whilst Delta is optimised for fusion at the cell surface, Omicron preferentially achieves entry through endosomal fusion; this biological about-face may impact transmission, cellular tropism and pathogenesis. Moreover, this switch away from TMPRSS2-mediated activation offers a mechanistic explanation for reduced syncytia formation by Omicron infected cells.
Discussion
The Omicron variant represents a major change in biological function and antigenicity of SARS-CoV-2. In this study, we demonstrate substantial immune escape of the Omicron variant. We present clear evidence of vaccine failure in dual-vaccinated individuals and partial restoration of immunity following a third booster dose of mRNA vaccine. In addition, we demonstrate a shift in the SARS-CoV-2 entry pathway from cell surface fusion, triggered by TMPRSS2, to cathepsin-dependent fusion within the endosome. This fundamental biological shift may affect the pathogenesis and severity of disease and requires further evaluation in population-based studies.
Using sera from double vaccine recipients, Omicron was found to be associated with a drop in neutralisation greater in magnitude than that reported in all other variants of concern (including Beta and Delta). Boosting enhanced neutralising responses to both the vaccine strain (Wuhan) and Omicron, particularly in recipients of ChAdOx1, but did not completely overcome the inherent immune escape properties of Omicron. Importantly, we did not assess the impact of vaccination on clinical severity of disease which is likely to be much higher than detection of infection. Protection against severe disease is longer lasting than prevention of infection. We also did not measure the impact of vaccination on T cell immunity which may be better preserved as only 14% of CD8+ and 28% of CD4+ epitopes are predicted to be affected by key Omicron mutations12.
The probability of infection with Omicron versus infection with the preceding Delta variant was significantly higher in double vaccine recipients, in keeping with the neutralisation data. A third dose of mRNA vaccine substantially reduced the probability of infection but did not restore immunity fully.
The emergence of a highly transmissible variant that is associated with escape from vaccine-induced immune responses means that over time, Omicron-specific vaccines may be required if disease severity was high, either directed at the general population or vulnerable groups. Early indications in young people are that Omicron infection is 40-70% less severe than Delta infection65, 66 similar calculations in the most vulnerable part of the population over the age of 40 years are awaited.
Genotypic changes in new variants have previously been shown to alter viral phenotype by modulating innate immune responses as well as evasion of the adaptive immune response67, 68. Additionally, mutations can alter spike functionality to impact transmission and pathogenesis24. Such changes may have provided emergent viruses with a selective advantage in lung cells and primary human airway epithelial cells. Enhanced spike activation by the plasma membrane protease TMPRSS2, may have enabled more rapid cell surface fusion54. In this study, we found that the Omicron variant has switched entry pathway to use TMPRSS2-independent endosomal fusion preferentially, a major change in the biological behaviour of the virus. This switching in the mechanism of fusion activation also manifests in reduced syncytia formation in infected cells, likely to reduce the cell-to-cell transmission characteristics of other variants. These properties have the potential to change cellular tropism and disease pathogenesis significantly. However, even a variant that is less virulent with a very high transmission rate may continue to present a substantial risk to older people and those with co-morbidities, especially those with immunosuppression. Moreover, our work demonstrates that SARS-CoV-2 exhibits high antigenic and functional plasticity; further fundamental shifts in transmission and disease should be anticipated.
Methods
Cells
Calu-3 are human lung adenocarcinoma epithelial cells. Caco-2 are an immortalized cell line derived from human colorectal adenocarcinoma, primarily used as a model of the intestinal epithelial barrier. A549 cells, a human alveolar adenocarcinoma line, were modified to stably express human ACE-2 and TMPRSS2. Human embryonic kidney (HEK293T) cells were used in pseudotype production. Baby Hamster Kidney clone 21 cells and Vero ACE-2 TMPRSS2 cells were used in the isolation of live Omicron SARS-CoV-2. All cell lines were maintained at 37°C and 5% CO2 in DMEM supplemented with 10% foetal bovine serum (FBS), except for Calu-3 cells which were supplemented with 20% FBS.
Generation of cell line expressing human ACE2 receptor
Lentiviral vectors encoding human ACE2 (GenBank NM_001371415.1) were produced as described previously69 and BHK-21 transduced cells were selected with 200µg/ml of hygromycin B.
Generation of cell lines used for fusion assays
Retrovirus vectors were produced by transfecting HEK-293T cells with plasmid pQCXIP-GFP1-10 (Addgene #68715) or pQCXIP-BSR-GFP11 (Addgene #68716)52 and packaging vectors expressing MLV gal-pol and VSV-G using Lipofectamine 3000 (Invitrogen) according to manufacturer’s instructions. Cell supernatants were harvested 24-48h post-transfection, pooled, clarified by centrifugation and filtered. One mL of each supernatant was used to transduce A549-Ace2-TMPRSS2 (AAT) cells69 in presence of Polybrene (Merck). Two days post-transduction, the supernatant was replaced with selection medium (DMEM 10% FBS 1µg/mL puromycin) and cells incubated until complete death of the untrasduced control cells were observed. The resulting puromycin-resistant cells (termed AAT-GFP1-10 and AAT-BSR-GFP11) were used in fusion assays.
Virus isolation from clinical samples
Nasopharyngeal swabs of patients infected with Omicron were collected with biorepository ethical approval (reference 20/ES/0061) in virus transport medium and resuspended in serum-free DMEM supplemented with 10 µg/ml gentamicin, 100 units/ml penicillin-streptomycin and 2.5µg/ml amphotericin B to a final volume of 1.5ml. Virus isolation was attempted in BHK-21 cells stably expressing the human ACE2 protein (BHK-hACE2) and VERO cells stably expressing ACE2 and TMPRSS2 (VAT69). The infected cells were incubated at 37°C and monitored for signs of cytopathic effect (CPE) and the presence of viral progeny in the medium by RT-qPCR. While no CPE was observed in any of the infected cells, RT-qPCR at 5 days post-infection (dpi) confirmed the presence of the virus derived from two of the five samples (referred to hereafter as 204 and 205) in the medium of BHK-hACE2, but not VAT cells (Extended data Fig. 7a). An aliquot of the clarified medium containing approximately 4×104 viral genomes of the P0 stocks of samples 204 and 205 was used to infect VAT, BHK-ACE2 and Calu-3 cells. No CPE was observed in the infected cells but once again, virus replication was confirmed in BHK-hACE2 and Calu-3 by RT-qPCR. Supernatants (termed P1) from infected Calu-3 cells at 3 dpi were collected and virus titrated by both focus forming assay and RT-qPCR. The virus reached more than 100-fold higher titres in Calu-3 cells compared to BHK-hACE2 (Extended data Fig. 7b). Further passage of sample 205-derived P1 virus in both Calu-3 and Caco-2 yielded equivalent genome copy numbers in both cell lines (Extended data Fig. 7b). CPE was observed at 3 dpi in both Calu-3 and Caco-2 cells (not shown). The medium (termed P2) of infected Calu-3 and Caco-2 cells was collected at 4 dpi, titrated and used in subsequent experiments.
Measurement of SARS-CoV-2, HCoVs and influenza antibody response by electrochemiluminescence
IgG antibody titres were measured quantitatively against SARS-CoV-2 trimeric spike (S) protein, N-terminal domain (NTD), receptor binding domain (RBD) or nucleocapsid (N), human seasonal coronaviruses (HCoVs) 229E, OC43, NL63 and HKU1; and influenza A (Michigan H1, Hong Kong H3 and Shanghai H7) and B (Phuket HA and Brisbane) using MSD V-PLEX COVID-19 Coronavirus Panel 2 (K15369) and Respiratory Panel 1 (K15365) kits. Multiplex Meso Scale Discovery electrochemiluminescence (MSD-ECL) assays were performed according to manufacturer instructions. Briefly, 96-well plates were blocked for one hour. Plates were then washed, samples were diluted 1:5000 in diluent and added to the plates along with serially diluted reference standard (calibrator) and serology controls 1.1, 1.2 and 1.3. After incubation, plates were washed and SULFO-TAG detection antibody added. Plates were washed and were immediately read using a MESO Sector S 600 plate reader. Data were generated by Methodological Mind software and analysed using MSD Discovery Workbench (v4.0). Results are expressed as MSD arbitrary units per ml (AU/ml). Reference plasma samples yielded the following values: negative pool - spike 56.6 AU/ml, NTD 119.4 AU/ml, RBD 110.5 AU/ml and nucleocapsid 20.7 AU/ml; SARS-CoV-2 positive pool - spike 1331.1 AU/ml, NTD 1545.2 AU/ml, RBD 1156.4 AU/ml and nucleocapsid 1549.0 AU/ml; NIBSC 20/130 reference - spike 547.7 AU/ml, NTD 538.8 AU/ml, RBD 536.9 AU/ml and nucleocapsid 1840.2 AU/ml.
Measurement of virus neutralising antibodies using viral pseudotypes
Pseudotype-based neutralisation assays were carried out as described previously 70,71,72. Briefly, HEK293, HEK293T, and 293-ACE2 71 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS, 200mM L-glutamine, 100µg/ml streptomycin and 100 IU/ml penicillin. HEK293T cells were transfected with the appropriate SARS-CoV-2 S gene expression vector (wild type or variant) in conjunction with p8.91 73 and pCSFLW 74 using polyethylenimine (PEI, Polysciences, Warrington, USA). HIV (SARS-CoV-2) pseudotypes containing supernatants were harvested 48 hours post-transfection, aliquoted and frozen at -80°C prior to use. S gene constructs bearing the WUHAN (D614G) and Omicron (B.1.1.529) S genes were based on the codon-optimised spike sequence of SARS-CoV-2 and generated by GeneArt (ThermoFisher). Constructs bore the following mutations relative to the Wuhan-Hu-1 sequence (GenBank: MN908947): WUHAN (D614G) – D614G; Omicron (BA.1, B.1.1.529) - A67V, Δ69-70, T95I, G142D/Δ143-145, Δ211/L212I, ins214EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F. 293-ACE2 target cells were maintained in complete DMEM supplemented with 2µg/ml puromycin.
Neutralising activity in each sample was measured by a serial dilution approach. Each sample was serially diluted in triplicate from 1:50 to 1:36450 in complete DMEM prior to incubation with HIV (SARS-CoV-2) pseudotypes, incubated for 1 hour, and plated onto 239-ACE2 target cells. After 48-72 hours, luciferase activity was quantified by the addition of Steadylite Plus chemiluminescence substrate and analysis on a Perkin Elmer EnSight multimode plate reader (Perkin Elmer, Beaconsfield, UK). Antibody titre was then estimated by interpolating the point at which infectivity had been reduced to 50% of the value for the no serum control samples.
Protease inhibitor studies
To selectively inhibit either cell surface or endosomal fusion of SARS-CoV-2, cells were pre-treated for one hour with 10µM of either Camostat mesylate (referred to hence forth as Camostat) or E64d prior to inoculation with pseudotype. In these studies, spike proteins from Alpha and Delta VOCs, and Guangdong isolate Pangolin coronavirus (GISAID ref EPI_ISL_410721) were used as controls.
Viral RNA extraction and RT-qPCR
Viral RNA was extracted from culture supernatants using the RNAdvance Blood kit (Beckman Coulter Life Sciences) following the manufacturer’s recommendations. RNA was used as template to detect and quantify viral genomes by duplex RT-qPCR using a Luna® Universal Probe One-Step RT-qPCR Kit (New England Biolabs, E3006E). SARS-CoV-2 specific RNAs were detected by targeting the N1 gene from the CDC panel as part of the SARS-CoV-2 Research Use Only qPCR Probe Kit (Integrated DNA Technologies) and the ORF1ab gene using the following set of primers and probes: SARS-CoV-2_Orf1ab_Forward 5’ GACATAGAAGTTACTGG&CGATAG 3’, SARS-CoV-2_Orf1ab_Reverse 5’ TTAATATGACGCGCACTACAG 3’, SARS-CoV-2_Orf1ab_Probe ACCCCGTGACCTTGGTGCTTGT with HEX/ZEN/3IABkFQ modifications. SARS-CoV-2 RNA was used to generate a standard curve and viral genomes were quantified and expressed as number of Orf1ab RNA molecules /ml of supernatant. All runs were performed on the ABI7500 Fast instrument and results analysed with the 7500 Software v2.3 (Applied Biosystems, Life Technologies).
Genome Sequencing and analysis
Sequencing was carried out by the UK public health agencies (UKHSA/PHE, PHS, PHW and PHNI) and by members of the COG-UK consortium using the ARTIC protocol as previously described. Sequences were aligned by mapping to the SARS-CoV-2 reference Wuhan-Hu-1 using Minimap2 75. Prior to phylogenetic analysis 85 sites exhibiting high genetic variability due to data quality issues in overseas sequencing labs were excluded using a masking script in Phylopipe (https://github.com/cov-ert/phylopipe). The phylogenetic tree was constructed with the maximum likelihood method FastTree2 76 using a JC+CAT nucleotide substitution model.
Replication curve
Calu-3 cells were seeded in a 96-well plate at a cell density of 3.5×10^4 cells per well. Cells were infected with the indicated viruses using the equivalent of 2×10^4 Orf1ab genome copies/well in serum-free RPMI-1640 medium (Gibco). After one hour of incubation at 37°C, cells were washed three times and left in 20% FBS RPMI-1640 medium. Supernatants were collected at different times post-infection and viral RNA extracted and quantified as described above.
Fusion assay
AAT-GFP1-10 and AAT-BSR-GFP11 cells were trypsinized and mixed at a ratio of 1:1 to seed a total of 2×10^4 cells/well in black 96-well plate (Greiner) in FluoroBrite DMEM medium (Thermo Fischer Scientific) supplemented with 2% FBS. Next day, cells were infected with the indicated viruses using the equivalent of 10^6 Orf1a genome copies/well in FluoroBrite DMEM 2% FBS. GFP signal was acquired for the following 20 hours using a CLARIOStar Plus (BMG LABTECH) equipped with ACU to maintain 37°C and 5% CO2. Data were analysed using MARS software and plotted with GraphPad prims 9 software. At 22 hs post-infection, cells were fixed in 8% formaldehyde, permeabilized with 0.1 % Triton X-100 and stained with sheep anti-SARS-CoV-2 N (1:500) antiserum 69 followed by Alexa Fluor 594 Donkey anti-sheep IgG (H+L) (1:500, Invitrogen) and DAPI (1:4000, Sigma). Cell imagines were acquired using EVOS Cell Imaging Systems (Thermo Fischer Scientific).
Demographic data
Data for the EVADE study were available using the NHS Greater Glasgow and Clyde (NHS GG&C) SafeHaven platform and included vaccination status (dates and product names for each dose), demographic data (age, sex and Scottish Index of Multiple Deprivation (SIMD) quartile) comorbidity (shielding and immunosuppression status) and dates of positive and negative PCR tests, for 1.2 million inhabitants of the (NHS GG&C) area over 18 years of age, from 1st March 2020 up to 12th January 2022. Data were matched by CHI number and pseudonymised before analysis. Derogated ethical approval was granted by the NHS GG&C SafeHaven committee (GSH/21/IM/001).
Vaccine effectiveness
We used a logistic additive regression model to estimate relative vaccine effectiveness against the Omicron variant as it emerged in a population of 1.2 million people in NHS Greater Glasgow & Clyde, the largest health board in Scotland. Infection status for Omicron and Delta was modelled by number and product type of vaccine doses, previous infection status, sex, SIMD quartile, age on 31st October 2021 and time since most recent vaccination.
We identified Omicron infections using 3 data streams: confirmed S gene target failure (SGTF), allele specific PCR, and Pango lineage assignments from the sequencing data. SGTF samples with Delta lineage assignments were assigned as Delta infections. Samples for which the sequencing date was more than two weeks away from the first positive PCR were removed from the analysis.
We removed a small number of individuals who received ChAdOx1 as a third dose or had their third dose before the first of September 2021 on the assumption that the majority were part of the COV-BOOST clinical trial, the results of which are published elsewhere. We removed anyone with ambiguous vaccination status or whose brand was unknown due to data entry error. We removed those who were vaccinated during the study period. We removed individuals who tested positive in the 90 days before the study period. To control for the effect of missed vaccinations due to recent infection, we exclude those who were eligible for a second or third dose but had not taken this up (i.e had a first dose more than 8 weeks ago but no second dose, or a second dose more than 12 weeks ago but no third dose). Since those who tested positive during the study period could not subsequently be vaccinated within the study period, and those who changed vaccination status during the study period were excluded from the dataset, there would be an inflated proportion of people testing positive and having only a second dose of a vaccine. Specifically, those who were given the third dose of a vaccine during the study period would be excluded from the analysis, while those who would have been given a third dose of the vaccine but could not due to becoming infected would be included in the analysis. This would lead to reduced estimates of effectiveness of second doses if not accounted for appropriately. Due to the timing of the rollout of booster doses coinciding with high levels of infection, it is vital to account for this.
Serum samples
Serum samples were collected from healthy participants in the COVID-19 Deployed Vaccine Cohort Study (DOVE), a cross-sectional post-licensing cohort study to determine the immunogenicity of deployed COVID-19 vaccines against evolving SARS-CoV-2 variants. 308 adult volunteers aged at least 18 years and were recruited into the study 14 days or more after a second or third dose of vaccine. All participants gave written informed consent to take part in the study. The DOVE study was approved by the North-West Liverpool Central Research Ethics Committee (REC reference 21/NW/0073).
Structural modelling
The file 6vsb_1_1_1.pdb containing a complete model of the full-length glycosylated spike homotrimer in open conformation with one monomer having the receptor-binding domain in the ‘up’ position was obtained from the CHARMM-GUI Archive 77, 78. This model is itself generated based upon a partial spike cryo-EM structure (PDB ID: 6VSB). For visualisation, the model was trimmed to the ectodomain (residues 14-1164) and the signal peptide (residues 1-13) and glycans were removed. Using this structural model and the closed conformation equivalent (6vxx_1_1_1.pdb). Residues belonging to the receptor-binding site were identified as those with an atom within 4Å of an ACE2 atom in the bound RBD-ACE2 structure (PDB ID: 6M0J79) and Alpha carbon-to-Alpha carbon distances between these residues in the ‘up’ RBD and all other spike residues were calculated. Antibody accessibility scores for open and closed conformations were calculated using BEpro 31. Figures were prepared using PyMol 80.
Epidemiological description of the emergence of the Omicron variant in the UK
On the 27th November 2021, the UK Health Security Agency detected 2 cases of Omicron in England, the following day 6 Scottish cases were detected by community (Pillar 2) sequencing. Over the next 10 days (to 8th December 2021) a further 95 genome sequences were obtained. Due to the rapid spread of Omicron and low genetic diversity, the genome sequences are highly related with mean genetic divergence of 1 single nucleotide polymorphisms (SNPs) and maximum 7 SNPs.
The phylogenetic relationship to Omicron sequences from other countries is consistent with multiple introductions associated with travel to South Africa followed by community transmissions within Scotland. Amongst the Scottish samples diverged from the tree backbone, there were a number identified that are genetically divergent, i.e., greater than 2 single nucleotide polymorphisms from the nearest Scottish sample (Fig. 1d). Moreover, comparison to the wider international collection of Omicron samples revealed that they were more closely related to genomes from other countries than other Scottish samples. These samples therefore likely represent independent introductions to Scotland, but without more detailed epidemiological data, the number of introductions is unknown. Where there are indistinguishable samples in the phylogeny from Scotland and elsewhere in world, importation cannot be ruled out as a source of these samples in Scotland, rather than transmission from an established population circulating in Scotland.
Within Scotland, cases are spread across 9 separate Health Boards and distributed throughout the phylogeny (Fig. 1d). Basal Scottish genomes were sampled in 7 different Health Boards, most of them from NHS Greater Glasgow & Clyde (47%) and NHS Lanarkshire (25%). Notably, amongst these earliest samples are cases that were epidemiologically linked to early spreading events. All but one of these samples were found on this basal branch and are indistinguishable, and which is consistent with transmission at these events.
Data and materials availability
The experimental data that support the findings of this study are available on reasonable request but restrictions apply to the availability of clinical data, which were used under ethical approvals for the current study, and so are not publicly available.
Codes used in this analysis are available in the study’s GitHub repository: https://github.com/centre-for-virus-research/Omicron.
Funding
Health Data Research UK (HDR UK) - the Evaluation of Variants Affecting Deployed COVID-19 Vaccine (EVADE) study (ECT, SR, OM, CW, BJW) (grant code: 2021.0155).
COG-UK is supported by funding from the Medical Research Council (MRC) part of UK Research & Innovation (UKRI), the National Institute of Health Research (NIHR) [grant code: MC_PC_19027], and Genome Research Limited, operating as the Wellcome Sanger Institute (RMB, DLR, ECT).
Medical Research Council (MRC) - the COVID-19 DeplOyed VaccinE (DOVE) study (grant code: MCUU1201412) and COG-UK (ECT).
Medical Research Council (MRC) (grant code: MC_UU_12014/12) (AF, JH, RO, JG, ECT and DLR)
Medical Research Council (MRC) (grant codes: MR/R024758/1 and MR/W005611/1) (WTH)
UK Research and Innovation (UKRI) - G2P-UK National Virology Consortium (MR/W005611/1) (MP, ECT, AP, DLR).
Wellcome Trust (grant code: 220977/Z/20/Z) (DLR)
Biotechnology and Biological Sciences Research Council (grant code: BBSRC, BB/R004250/1) (NL, BJW)
Wellcome Trust and Royal Society Sir Henry Dale Fellowship (grant code: 107653/Z/15/A) (JG)
Author contributions
Conceptualization: ECT, JG, MP, DLR, BW
Methodology: BJW, JG, OAM, CW, NL, GL, WF, SS, MM, AS, WTH, CD, RO, JH, DP, KP, AF, GY, SS, RMP, PRM, AHP, JH, DLR, SR
Investigation: BJW, JG, OAM, CW, NL, GL, WF, SS, MM, AS, SA, EV, WTH, CD, RO, JH, PH, VS, DP, KP, AF, GY, SS, MTGH, RG, KT,AHP, JH,DLR,MP,SR
Visualization: BJW, JG, OAM, CW, GL, WTH, DLR
Funding acquisition: ECT, DLR, SR, BJW, JG
Project administration: ECT, DLR, SR, JH,
Supervision: BJW, DLR, MP, SR, JH, ECT
Writing – original draft: ECT, BJW, JG, GY
Writing – review & editing: BJW, JG, OAM, CW, SA, WTH, DLR, MP, ECT
Competing interests
The authors declare that they have no competing interests.
Data and materials availability
The experimental data that support the findings of this study are available on reasonable request but restrictions apply to the availability of clinical data, which were used under ethical approvals for the current study, and so are not publicly available. Anonymised data are available with permission of the NHS Greater Glasgow & Clyde SafeHaven.
Codes used in this analysis are available upon request from the corresponding author.
Biological materials including cell lines are available on reasonable request from the authors. Clinical samples are restricted for use under the ethical approvals obtained for their use.
Author information
These authors contributed equally: Brian J. Willett, Joe Grove, Oscar A. MacLean, Craig Wilkie, Nicola Logan, Giuditta De Lorenzo & Wilhelm Furnon
These authors contributed equally: John Haughney, David L. Robertson, Massimo Palmarini, Surajit Ray & Emma C. Thomson
Contributions
Conceptualization: ECT, JG, MP, DLR, BW
Methodology: BJW, JG, OAM, CW, NL, GL, WF, SS, MM, AS, WTH, CD, RO, JH, DP, KP, AF, GY, SS, RMP, PRM, AHP, JH, DLR, SR
Investigation: BJW, JG, OAM, CW, NL, GL, WF, SS, MM, AS, SA, EV, WTH, CD, RO, JH, PH, VS, DP, KP, AF, GY, SS, MTGH, RG, KT, AHP, JH, DLR, MP, SR
Visualization: BJW, JG, OAM, CW, GL, WTH, DLR
Funding acquisition: ECT, DLR, SR, BJW, JG
Project administration: ECT, DLR, SR, JH
Supervision: BJW, DLR, MP, SR, JH, ECT
Writing – original draft: ECT, BJW, JG, GY
Writing – review & editing: BJW, JG, OAM, CW, SA, WTH, DLR, MP, ECT
Corresponding authors
Correspondence to Professor Emma Thomson (emma.thomson{at}glasgow.ac.uk) and Professor Brian Willett (brian.willett{at}glasgow.ac.uk), MRC-University of Glasgow Centre for Virus Research, UK, G61 1QH.
Ethics declarations
The authors declare that they have no competing interests.
Extended Data
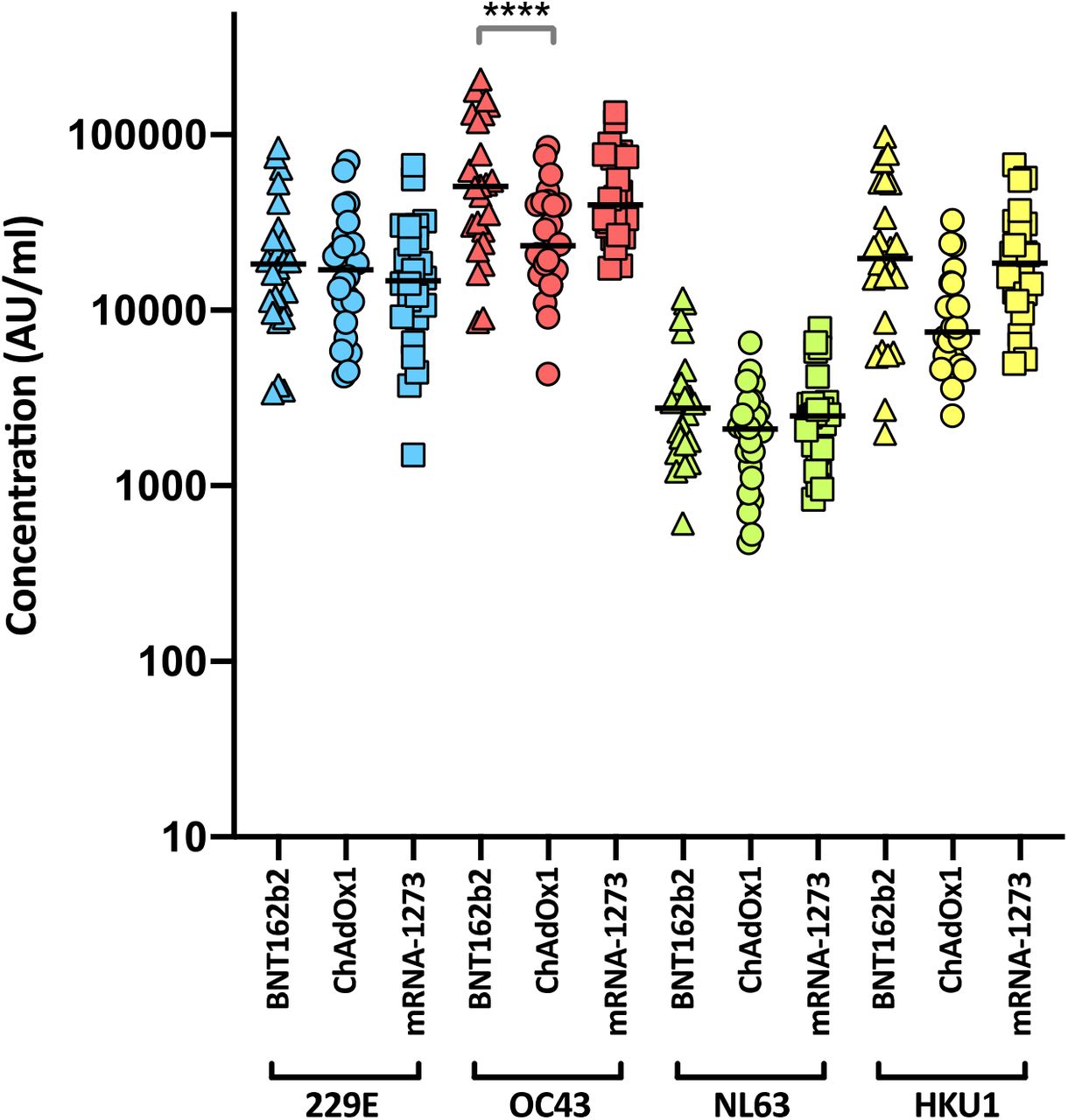
Antibody responses were studied in three groups of individuals (n=24 per group) vaccinated with either BNT162b2, ChAdOx1 or mRNA-1273 by MSD-ECL assay. Responses were measured against full-length spike glycoprotein (Spike) from HCoVs 229E, OC43, NL63 and HKU1 and are expressed as MSD arbitrary units (AU/ml). The response to OC43 was significantly higher in BNT162b2 vaccinates than in ChAdOx1 vaccinates.

Antibody responses were studied in three groups of individuals (n=24 per group) vaccinated with either BNT162b2, ChAdOx1 or mRNA-1273 by MSD-ECL assay. Responses were measured against haemagglutinins from influenza viruses; influenza A Michigan H1, Hong Kong H3 and Shanghai H7, and influenza B Phuket HA and Brisbane and are expressed as MSD arbitrary units (AU/ml). No significant differences were detected between the vaccine groups for each of the antigens.

Antibody responses were studied in four groups of individuals primed with two doses of either ChAdOx1 or BNT162b2, followed by a booster of BNT162b2 or mRNA-1273. Responses were measured by MSD-ECL assay against full-length spike glycoprotein (Spike) from HCoVs 229E, OC43, NL63 and HKU1 and are expressed as MSD arbitrary units (AU/ml).
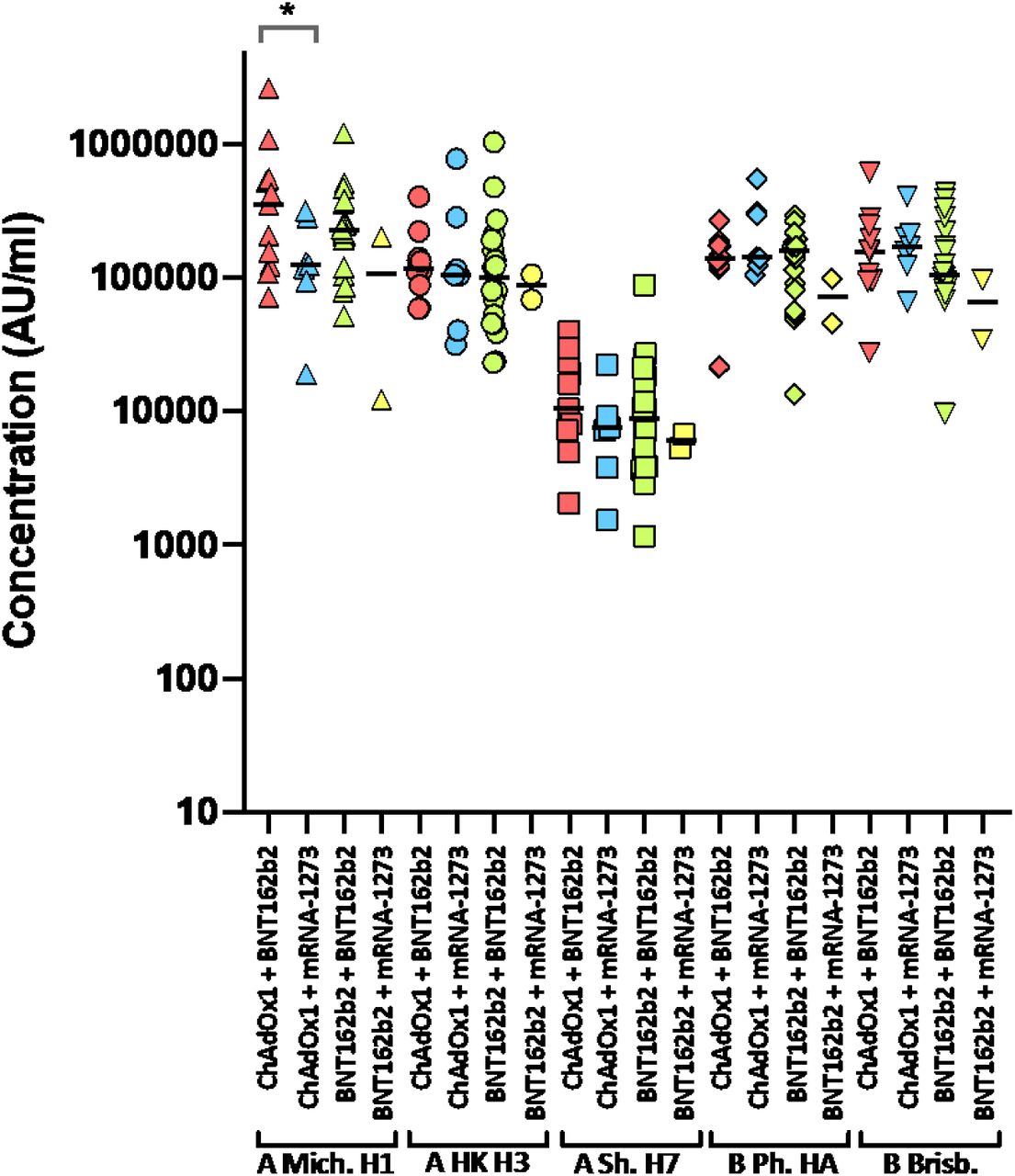
Antibody responses were studied in four groups of individuals primed with two doses of either ChAdOx1 or BNT162b2, followed by a booster of BNT162b2 or mRNA-1273. Responses were measured by MSD-ECL against haemagglutinins from influenza viruses; influenza A Michigan H1, Hong Kong H3 and Shanghai H7, and influenza B Phuket HA and Brisbane and are expressed as MSD arbitrary units (AU/ml). * Significantly different p=0.0413.

Two groups of healthy volunteers vaccinated with two doses of either ChAdOx1 or BNT162b2, were sampled two weeks following a third dose of either BNT162b2 or mRNA-1273. Each point represents the mean of three replicates. Where dose 2 and dose 3 samples were available from the same individual, points are joined by a solid line.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
a, Vero ACE2 TMPRSS2 (VAT) and BHK-hACE2 cells were inoculated with diluted clinical samples. Viral progeny was quantified in the medium 5 dpi by RT-qPCR. b, Aliquots of the medium from samples named 204 and 205 were used to generate a P1 in BHK-hACE2 and Calu-3 cells and, limited to sample 205, a P2 in Calu-3 and Caco2 cells. Viral stocks were quantified by RT-qPCR.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA.
Neutralising antibody responses were quantified against Wuhan or Omicron spike glycoprotein-bearing HIV(SARS-CoV-2) pseudotypes. Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA. P= BNT162b2, AZ = ChAdOx1, M = mRNA-1273.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA. P= BNT162b2, AZ = ChAdOx1, M = mRNA-1273.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA. P= BNT162b2, AZ = ChAdOx1, M = mRNA-1273.
Data were analyzed in GraphPad Prism v8.4.3, groups were compared by ordinary one-way ANOVA.
Population consisted of 101,310 people aged 18 and over, registered as living in NHS Greater Glasgow and Clyde and tested by PCR test for SARS-CoV-2 infection between 22nd and 28th December 2021, split by SARS-CoV-2 variant status.
The COVID-19 Genomics UK (COG-UK) consortium - June 2021 V.1
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Samuel C Robson 13, 84
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr Thomas R Connor 11, 74 and Prof Nicholas J Loman 43
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Tanya Golubchik 5
Funding acquisition, Leadership and supervision, Metadata curation, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Rocio T Martinez Nunez 46
Funding acquisition, Leadership and supervision, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr David Bonsall 5
Funding acquisition, Leadership and supervision, Project administration, Sequencing and analysis, Software and analysis tools, and Visualisation:
Prof Andrew Rambaut 104
Funding acquisition, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr Luke B Snell 12
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Software and analysis tools, and Visualisation:
Rich Livett 116
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Dr Catherine Ludden 20, 70
Funding acquisition, Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Dr Sally Corden 74 and Dr Eleni Nastouli 96, 95, 30
Funding acquisition, Leadership and supervision, Metadata curation, Sequencing and analysis, and Software and analysis tools:
Dr Gaia Nebbia 12
Funding acquisition, Leadership and supervision, Project administration, Samples and logistics, and Sequencing and analysis:
Ian Johnston 116
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
Prof Katrina Lythgoe 5, Dr M. Estee Torok 19, 20 and Prof Ian G Goodfellow 24
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Visualisation:
Dr Jacqui A Prieto 97, 82 and Dr Kordo Saeed 97, 83
Leadership and supervision, Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
Dr David K Jackson 116
Leadership and supervision, Metadata curation, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Catherine Houlihan 96, 94
Leadership and supervision, Metadata curation, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Dan Frampton 94, 95
Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr William L Hamilton 19 and Dr Adam A Witney 41
Funding acquisition, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Giselda Bucca 101
Funding acquisition, Leadership and supervision, Metadata curation, and Project administration:
Dr Cassie F Pope 40, 41
Funding acquisition, Leadership and supervision, Metadata curation, and Samples and logistics:
Dr Catherine Moore 74
Funding acquisition, Leadership and supervision, Metadata curation, and Sequencing and analysis:
Prof Emma C Thomson 53
Funding acquisition, Leadership and supervision, Project administration, and Samples and logistics:
Dr Ewan M Harrison 116, 102
Funding acquisition, Leadership and supervision, Sequencing and analysis, and Visualisation:
Prof Colin P Smith 101
Leadership and supervision, Metadata curation, Project administration, and Sequencing and analysis:
Fiona Rogan 77
Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Shaun M Beckwith 6, Abigail Murray 6, Dawn Singleton 6, Dr Kirstine Eastick 37, Dr Liz A Sheridan 98, Paul Randell 99, Dr Leigh M Jackson 105, Dr Cristina V Ariani 116 and Dr Sónia Gonçalves 116
Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Dr Derek J Fairley 3, 77, Prof Matthew W Loose 18 and Joanne Watkins 74
Leadership and supervision, Metadata curation, Samples and logistics, and Visualisation:
Dr Samuel Moses 25, 106
Leadership and supervision, Metadata curation, Sequencing and analysis, and Software and analysis tools:
Dr Sam Nicholls 43, Dr Matthew Bull 74 and Dr Roberto Amato 116
Leadership and supervision, Project administration, Samples and logistics, and Sequencing and analysis:
Prof Darren L Smith 36, 65, 66
Leadership and supervision, Sequencing and analysis, Software and analysis tools, and Visualisation:
Prof David M Aanensen 14, 116 and Dr Jeffrey C Barrett 116
Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
Dr Dinesh Aggarwal 20, 116, 70, Dr James G Shepherd 53, Dr Martin D Curran 71 and Dr Surendra Parmar71
Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
Dr Matthew D Parker 109
Metadata curation, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr Catryn Williams 74
Metadata curation, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Sharon Glaysher 68
Metadata curation, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Anthony P Underwood 14, 116, Dr Matthew Bashton 36, 65, Dr Nicole Pacchiarini 74, Dr Katie F Loveson 84 and Matthew Byott 95, 96
Project administration, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Alessandro M Carabelli 20
Funding acquisition, Leadership and supervision, and Metadata curation:
Dr Kate E Templeton 56, 104
Funding acquisition, Leadership and supervision, and Project administration:
Dr Thushan I de Silva 109, Dr Dennis Wang 109, Dr Cordelia F Langford 116 and John Sillitoe 116
Funding acquisition, Leadership and supervision, and Samples and logistics:
Prof Rory N Gunson 55
Funding acquisition, Leadership and supervision, and Sequencing and analysis:
Dr Simon Cottrell 74, Dr Justin O’Grady 75, 103 and Prof Dominic Kwiatkowski 116, 108
Leadership and supervision, Metadata curation, and Project administration:
Dr Patrick J Lillie 37
Leadership and supervision, Metadata curation, and Samples and logistics:
Dr Nicholas Cortes 33, Dr Nathan Moore 33, Dr Claire Thomas 33, Phillipa J Burns 37, Dr Tabitha W Mahungu 80 and Steven Liggett 86
Leadership and supervision, Metadata curation, and Sequencing and analysis:
Angela H Beckett 13, 81 and Prof Matthew TG Holden 73
Leadership and supervision, Project administration, and Samples and logistics:
Dr Lisa J Levett 34, Dr Husam Osman 70, 35 and Dr Mohammed O Hassan-Ibrahim 99
Leadership and supervision, Project administration, and Sequencing and analysis:
Dr David A Simpson 77
Leadership and supervision, Samples and logistics, and Sequencing and analysis:
Dr Meera Chand 72, Prof Ravi K Gupta 102, Prof Alistair C Darby 107 and Prof Steve Paterson 107
Leadership and supervision, Sequencing and analysis, and Software and analysis tools:
Prof Oliver G Pybus 23, Dr Erik M Volz 39, Prof Daniela de Angelis 52, Prof David L Robertson 53, Dr Andrew J Page 75 and Dr Inigo Martincorena 116
Leadership and supervision, Sequencing and analysis, and Visualisation:
Dr Louise Aigrain 116 and Dr Andrew R Bassett 116
Metadata curation, Project administration, and Samples and logistics:
Dr Nick Wong 50, Dr Yusri Taha 89, Michelle J Erkiert 99 and Dr Michael H Spencer Chapman 116, 102
Metadata curation, Project administration, and Sequencing and analysis:
Dr Rebecca Dewar 56 and Martin P McHugh 56, 111
Metadata curation, Project administration, and Software and analysis tools:
Siddharth Mookerjee 38, 57
Metadata curation, Project administration, and Visualisation:
Stephen Aplin 97, Matthew Harvey 97, Thea Sass 97, Dr Helen Umpleby 97 and Helen Wheeler 97
Metadata curation, Samples and logistics, and Sequencing and analysis:
Dr James P McKenna 3, Dr Ben Warne 9, Joshua F Taylor 22, Yasmin Chaudhry 24, Rhys Izuagbe 24, Dr Aminu S Jahun 24, Dr Gregory R Young 36, 65, Dr Claire McMurray 43, Dr Clare M McCann 65, 66, Dr Andrew Nelson 65, 66 and Scott Elliott 68
Metadata curation, Samples and logistics, and Visualisation:
Hannah Lowe 25
Metadata curation, Sequencing and analysis, and Software and analysis tools:
Dr Anna Price 11, Matthew R Crown 65, Dr Sara Rey 74, Dr Sunando Roy 96 and Dr Ben Temperton 105
Metadata curation, Sequencing and analysis, and Visualisation:
Dr Sharif Shaaban 73 and Dr Andrew R Hesketh 101
Project administration, Samples and logistics, and Sequencing and analysis:
Dr Kenneth G Laing 41, Dr Irene M Monahan 41 and Dr Judith Heaney 95, 96, 34
Project administration, Samples and logistics, and Visualisation:
Dr Emanuela Pelosi 97, Siona Silviera 97 and Dr Eleri Wilson-Davies 97
Samples and logistics, Software and analysis tools, and Visualisation:
Dr Helen Fryer 5
Sequencing and analysis, Software and analysis tools, and Visualization:
Dr Helen Adams 4, Dr Louis du Plessis 23, Dr Rob Johnson 39, Dr William T Harvey 53, 42, Dr Joseph Hughes 53, Dr Richard J Orton 53, Dr Lewis G Spurgin 59, Dr Yann Bourgeois 81, Dr Chris Ruis 102, Áine O’Toole 104, Marina Gourtovaia 116 and Dr Theo Sanderson 116
Funding acquisition, and Leadership and supervision:
Dr Christophe Fraser 5, Dr Jonathan Edgeworth 12, Prof Judith Breuer 96, 29, Dr Stephen L Michell 105 and Prof John A Todd 115
Funding acquisition, and Project administration:
Michaela John 10 and Dr David Buck 115
Leadership and supervision, and Metadata curation:
Dr Kavitha Gajee 37 and Dr Gemma L Kay 75
Leadership and supervision, and Project administration:
Prof Sharon J Peacock 20, 70 and David Heyburn 74
Leadership and supervision, and Samples and logistics:
Katie Kitchman 37, Prof Alan McNally 43, 93, David T Pritchard 50, Dr Samir Dervisevic 58, Dr Peter Muir70, Dr Esther Robinson 70, 35, Dr Barry B Vipond 70, Newara A Ramadan 78, Dr Christopher Jeanes 90, Danni Weldon 116, Jana Catalan 118 and Neil Jones 118
Leadership and supervision, and Sequencing and analysis:
Dr Ana da Silva Filipe 53, Dr Chris Williams 74, Marc Fuchs 77, Dr Julia Miskelly 77, Dr Aaron R Jeffries 105, Karen Oliver 116 and Dr Naomi R Park 116
Metadata curation, and Samples and logistics:
Amy Ash 1, Cherian Koshy 1, Magdalena Barrow 7, Dr Sarah L Buchan 7, Dr Anna Mantzouratou 7, Dr Gemma Clark 15, Dr Christopher W Holmes 16, Sharon Campbell 17, Thomas Davis 21, Ngee Keong Tan 22, Dr Julianne R Brown 29, Dr Kathryn A Harris 29, 2, Stephen P Kidd 33, Dr Paul R Grant 34, Dr Li Xu- McCrae 35, Dr Alison Cox 38, 63, Pinglawathee Madona 38, 63, Dr Marcus Pond 38, 63, Dr Paul A Randell 38, 63, Karen T Withell 48, Cheryl Williams 51, Dr Clive Graham 60, Rebecca Denton-Smith 62, Emma Swindells 62, Robyn Turnbull 62, Dr Tim J Sloan 67, Dr Andrew Bosworth 70, 35, Stephanie Hutchings 70, Hannah M Pymont 70, Dr Anna Casey 76, Dr Liz Ratcliffe 76, Dr Christopher R Jones 79, 105, Dr Bridget A Knight 79, 105, Dr Tanzina Haque 80, Dr Jennifer Hart 80, Dr Dianne Irish-Tavares 80, Eric Witele 80, Craig Mower 86, Louisa K Watson 86, Jennifer Collins 89, Gary Eltringham 89, Dorian Crudgington 98, Ben Macklin 98, Prof Miren Iturriza-Gomara 107, Dr Anita O Lucaci 107 and Dr Patrick C McClure 113
Metadata curation, and Sequencing and analysis:
Matthew Carlile 18, Dr Nadine Holmes 18, Dr Christopher Moore 18, Dr Nathaniel Storey 29, Dr Stefan Rooke 73, Dr Gonzalo Yebra 73, Dr Noel Craine 74, Malorie Perry 74, Dr Nabil-Fareed Alikhan 75, Dr Stephen Bridgett 77, Kate F Cook 84, Christopher Fearn 84, Dr Salman Goudarzi 84, Prof Ronan A Lyons 88, Dr Thomas Williams 104, Dr Sam T Haldenby 107, Jillian Durham 116 and Dr Steven Leonard 116
Metadata curation, and Software and analysis tools:
Robert M Davies 116
Project administration, and Samples and logistics:
Dr Rahul Batra 12, Beth Blane 20, Dr Moira J Spyer 30, 95, 96, Perminder Smith 32, 112, Mehmet Yavus 85, 109, Dr Rachel J Williams 96, Dr Adhyana IK Mahanama 97, Dr Buddhini Samaraweera 97, Sophia T Girgis 102, Samantha E Hansford 109, Dr Angie Green 115, Dr Charlotte Beaver 116, Katherine L Bellis 116, 102, Matthew J Dorman 116, Sally Kay 116, Liam Prestwood 116 and Dr Shavanthi Rajatileka 116
Project administration, and Sequencing and analysis:
Dr Joshua Quick 43
Project administration, and Software and analysis tools:
Radoslaw Poplawski 43
Samples and logistics, and Sequencing and analysis:
Dr Nicola Reynolds 8, Andrew Mack 11, Dr Arthur Morriss 11, Thomas Whalley 11, Bindi Patel 12, Dr Iliana Georgana 24, Dr Myra Hosmillo 24, Malte L Pinckert 24, Dr Joanne Stockton 43, Dr John H Henderson 65, Amy Hollis 65, Dr William Stanley 65, Dr Wen C Yew 65, Dr Richard Myers 72, Dr Alicia Thornton 72, Alexander Adams 74, Tara Annett 74, Dr Hibo Asad 74, Alec Birchley 74, Jason Coombes 74, Johnathan M Evans 74, Laia Fina 74, Bree Gatica-Wilcox 74, Lauren Gilbert 74, Lee Graham 74, Jessica Hey 74, Ember Hilvers 74, Sophie Jones 74, Hannah Jones 74, Sara Kumziene-Summerhayes 74, Dr Caoimhe McKerr 74, Jessica Powell 74, Georgia Pugh 74, Sarah Taylor 74, Alexander J Trotter 75, Charlotte A Williams 96, Leanne M Kermack 102, Benjamin H Foulkes 109, Marta Gallis 109, Hailey R Hornsby 109, Stavroula F Louka 109, Dr Manoj Pohare 109, Paige Wolverson 109, Peijun Zhang 109, George MacIntyre-Cockett 115, Amy Trebes 115, Dr Robin J Moll 116, Lynne Ferguson 117, Dr Emily J Goldstein 117, Dr Alasdair Maclean 117 and Dr Rachael Tomb 117
Samples and logistics, and Software and analysis tools:
Dr Igor Starinskij 53
Sequencing and analysis, and Software and analysis tools:
Laura Thomson 5, Joel Southgate 11, 74, Dr Moritz UG Kraemer 23, Dr Jayna Raghwani 23, Dr Alex E Zarebski 23, Olivia Boyd 39, Lily Geidelberg 39, Dr Chris J Illingworth 52, Dr Chris Jackson 52, Dr David Pascall 52, Dr Sreenu Vattipally 53, Timothy M Freeman 109, Dr Sharon N Hsu 109, Dr Benjamin B Lindsey109, Dr Keith James 116, Kevin Lewis 116, Gerry Tonkin-Hill 116 and Dr Jaime M Tovar-Corona 116
Sequencing and analysis, and Visualisation:
MacGregor Cox 20
Software and analysis tools, and Visualisation:
Dr Khalil Abudahab 14, 116, Mirko Menegazzo 14, Ben EW Taylor MEng 14, 116, Dr Corin A Yeats 14, Afrida Mukaddas 53, Derek W Wright 53, Dr Leonardo de Oliveira Martins 75, Dr Rachel Colquhoun 104, Verity Hill 104, Dr Ben Jackson 104, Dr JT McCrone 104, Dr Nathan Medd 104, Dr Emily Scher 104 and Jon-Paul Keatley 116
Leadership and supervision:
Dr Tanya Curran 3, Dr Sian Morgan 10, Prof Patrick Maxwell 20, Prof Ken Smith 20, Dr Sahar Eldirdiri 21, Anita Kenyon 21, Prof Alison H Holmes 38, 57, Dr James R Price 38, 57, Dr Tim Wyatt 69, Dr Alison E Mather 75, Dr Timofey Skvortsov 77 and Prof John A Hartley 96
Metadata curation:
Prof Martyn Guest 11, Dr Christine Kitchen 11, Dr Ian Merrick 11, Robert Munn 11, Dr Beatrice Bertolusso 33, Dr Jessica Lynch 33, Dr Gabrielle Vernet 33, Stuart Kirk 34, Dr Elizabeth Wastnedge 56, Dr Rachael Stanley 58, Giles Idle 64, Dr Declan T Bradley 69, 77, Dr Jennifer Poyner 79 and Matilde Mori 110
Project administration:
Owen Jones 11, Victoria Wright 18, Ellena Brooks 20, Carol M Churcher 20, Mireille Fragakis 20, Dr Katerina Galai 20, 70, Dr Andrew Jermy 20, Sarah Judges 20, Georgina M McManus 20, Kim S Smith 20, Dr Elaine Westwick 20, Dr Stephen W Attwood 23, Dr Frances Bolt 38, 57, Dr Alisha Davies 74, Elen De Lacy 74, Fatima Downing 74, Sue Edwards 74, Lizzie Meadows 75, Sarah Jeremiah 97, Dr Nikki Smith 109 and Luke Foulser 116
Samples and logistics:
Dr Themoula Charalampous 12, 46, Amita Patel 12, Dr Louise Berry 15, Dr Tim Boswell 15, Dr Vicki M Fleming 15, Dr Hannah C Howson-Wells 15, Dr Amelia Joseph 15, Manjinder Khakh 15, Dr Michelle M Lister 15, Paul W Bird 16, Karlie Fallon 16, Thomas Helmer 16, Dr Claire L McMurray 16, Mina Odedra 16, Jessica Shaw 16, Dr Julian W Tang 16, Nicholas J Willford 16, Victoria Blakey 17, Dr Veena Raviprakash 17, Nicola Sheriff 17, Lesley-Anne Williams 17, Theresa Feltwell 20, Dr Luke Bedford 26, Dr James S Cargill 27, Warwick Hughes 27, Dr Jonathan Moore 28, Susanne Stonehouse 28, Laura Atkinson 29, Jack CD Lee 29, Dr Divya Shah 29, Adela Alcolea-Medina 32, 112, Natasha Ohemeng-Kumi 32, 112, John Ramble 32, 112, Jasveen Sehmi 32, 112, Dr Rebecca Williams 33, Wendy Chatterton 34, Monika Pusok 34, William Everson 37, Anibolina Castigador 44, Emily Macnaughton 44, Dr Kate El Bouzidi 45, Dr Temi Lampejo 45, Dr Malur Sudhanva 45, Cassie Breen 47, Dr Graciela Sluga 48, Dr Shazaad SY Ahmad 49, 70, Dr Ryan P George 49, Dr Nicholas W Machin 49, 70, Debbie Binns 50, Victoria James 50, Dr Rachel Blacow 55, Dr Lindsay Coupland 58, Dr Louise Smith 59, Dr Edward Barton 60, Debra Padgett 60, Garren Scott 60, Dr Aidan Cross 61, Dr Mariyam Mirfenderesky 61, Jane Greenaway 62, Kevin Cole 64, Phillip Clarke 67, Nichola Duckworth 67, Sarah Walsh 67, Kelly Bicknell 68, Robert Impey 68, Dr Sarah Wyllie 68, Richard Hopes 70, Dr Chloe Bishop 72, Dr Vicki Chalker 72, Dr Ian Harrison 72, Laura Gifford 74, Dr Zoltan Molnar 77, Dr Cressida Auckland 79, Dr Cariad Evans 85, 109, Dr Kate Johnson 85, 109, Dr David G Partridge 85, 109, Dr Mohammad Raza 85, 109, Paul Baker 86, Prof Stephen Bonner 86, Sarah Essex 86, Leanne J Murray 86, Andrew I Lawton 87, Dr Shirelle Burton-Fanning 89, Dr Brendan AI Payne 89, Dr Sheila Waugh 89, Andrea N Gomes 91, Maimuna Kimuli 91, Darren R Murray 91, Paula Ashfield 92, Dr Donald Dobie 92, Dr Fiona Ashford 93, Dr Angus Best 93, Dr Liam Crawford 93, Dr Nicola Cumley 93, Dr Megan Mayhew 93, Dr Oliver Megram 93, Dr Jeremy Mirza 93, Dr Emma Moles-Garcia 93, Dr Benita Percival 93, Megan Driscoll 96, Leah Ensell 96, Dr Helen L Lowe 96, Laurentiu Maftei 96, Matteo Mondani 96, Nicola J Chaloner 99, Benjamin J Cogger99, Lisa J Easton 99, Hannah Huckson 99, Jonathan Lewis 99, Sarah Lowdon 99, Cassandra S Malone 99, Florence Munemo 99, Manasa Mutingwende 99, Roberto Nicodemi 99, Olga Podplomyk 99, Thomas Somassa 99, Dr Andrew Beggs 100, Dr Alex Richter 100, Claire Cormie 102, Joana Dias 102, Sally Forrest 102, Dr Ellen E Higginson 102, Mailis Maes 102, Jamie Young 102, Dr Rose K Davidson 103, Kathryn A Jackson 107, Dr Lance Turtle 107, Dr Alexander J Keeley 109, Prof Jonathan Ball 113, Timothy Byaruhanga 113, Dr Joseph G Chappell 113, Jayasree Dey 113, Jack D Hill 113, Emily J Park 113, Arezou Fanaie 114, Rachel A Hilson 114, Geraldine Yaze 114 and Stephanie Lo 116
Sequencing and analysis:
Safiah Afifi 10, Robert Beer 10, Joshua Maksimovic 10, Kathryn McCluggage 10, Karla Spellman 10, Catherine Bresner 11, William Fuller 11, Dr Angela Marchbank 11, Trudy Workman 11, Dr Ekaterina Shelest 13, 81, Dr Johnny Debebe 18, Dr Fei Sang 18, Dr Marina Escalera Zamudio 23, Dr Sarah Francois 23, Bernardo Gutierrez 23, Dr Tetyana I Vasylyeva 23, Dr Flavia Flaviani 31, Dr Manon Ragonnet-Cronin 39, Dr Katherine L Smollett 42, Alice Broos 53, Daniel Mair 53, Jenna Nichols 53, Dr Kyriaki Nomikou 53, Dr Lily Tong 53, Ioulia Tsatsani 53, Prof Sarah O’Brien 54, Prof Steven Rushton 54, Dr Roy Sanderson 54, Dr Jon Perkins 55, Seb Cotton 56, Abbie Gallagher 56, Dr Elias Allara 70, 102, Clare Pearson 70, 102, Dr David Bibby 72, Dr Gavin Dabrera 72, Dr Nicholas Ellaby 72, Dr Eileen Gallagher 72, Dr Jonathan Hubb 72, Dr Angie Lackenby 72, Dr David Lee 72, Nikos Manesis 72, Dr Tamyo Mbisa 72, Dr Steven Platt 72, Katherine A Twohig 72, Dr Mari Morgan 74, Alp Aydin 75, David J Baker 75, Dr Ebenezer Foster-Nyarko 75, Dr Sophie J Prosolek 75, Steven Rudder 75, Chris Baxter 77, Sílvia F Carvalho 77, Dr Deborah Lavin 77, Dr Arun Mariappan 77, Dr Clara Radulescu 77, Dr Aditi Singh 77, Miao Tang 77, Helen Morcrette 79, Nadua Bayzid 96, Marius Cotic 96, Dr Carlos E Balcazar 104, Dr Michael D Gallagher 104, Dr Daniel Maloney 104, Thomas D Stanton 104, Dr Kathleen A Williamson 104, Dr Robin Manley 105, Michelle L Michelsen 105, Dr Christine M Sambles 105, Dr David J Studholme 105, Joanna Warwick-Dugdale 105, Richard Eccles 107, Matthew Gemmell 107, Dr Richard Gregory 107, Dr Margaret Hughes 107, Charlotte Nelson 107, Dr Lucille Rainbow 107, Dr Edith E Vamos 107, Hermione J Webster 107, Dr Mark Whitehead 107, Claudia Wierzbicki 107, Dr Adrienn Angyal 109, Dr Luke R Green 109, Dr Max Whiteley 109, Emma Betteridge 116, Dr Iraad F Bronner 116, Ben W Farr 116, Scott Goodwin 116, Dr Stefanie V Lensing 116, Shane A McCarthy 116, 102, Dr Michael A Quail 116, Diana Rajan 116, Dr Nicholas M Redshaw 116, Carol Scott 116, Lesley Shirley 116 and Scott AJ Thurston 116
Software and analysis tools:
Dr Will Rowe 43, Amy Gaskin 74, Dr Thanh Le-Viet 75, James Bonfield 116, Jennifier Liddle 116 and Andrew Whitwham 116
1 Barking, Havering and Redbridge University Hospitals NHS Trust, 2 Barts Health NHS Trust, 3 Belfast Health & Social Care Trust, 4 Betsi Cadwaladr University Health Board, 5 Big Data Institute, Nuffield Department of Medicine, University of Oxford, 6 Blackpool Teaching Hospitals NHS Foundation Trust, 7 Bournemouth University, 8 Cambridge Stem Cell Institute, University of Cambridge, 9 Cambridge University Hospitals NHS Foundation Trust, 10 Cardiff and Vale University Health Board, 11 Cardiff University, 12 Centre for Clinical Infection and Diagnostics Research, Department of Infectious Diseases, Guy’s and St Thomas’ NHS Foundation Trust, 13 Centre for Enzyme Innovation, University of Portsmouth, 14 Centre for Genomic Pathogen Surveillance, University of Oxford, 15 Clinical Microbiology Department, Queens Medical Centre, Nottingham University Hospitals NHS Trust, 16 Clinical Microbiology, University Hospitals of Leicester NHS Trust, 17 County Durham and Darlington NHS Foundation Trust, 18 Deep Seq, School of Life Sciences, Queens Medical Centre, University of Nottingham, 19 Department of Infectious Diseases and Microbiology, Cambridge University Hospitals NHS Foundation Trust, 20 Department of Medicine, University of Cambridge, 21 Department of Microbiology, Kettering General Hospital, 22 Department of Microbiology, South West London Pathology, 23 Department of Zoology, University of Oxford, 24 Division of Virology, Department of Pathology, University of Cambridge, 25 East Kent Hospitals University NHS Foundation Trust, 26 East Suffolk and North Essex NHS Foundation Trust, 27 East Sussex Healthcare NHS Trust, 28 Gateshead Health NHS Foundation Trust, 29 Great Ormond Street Hospital for Children NHS Foundation Trust, 30 Great Ormond Street Institute of Child Health (GOS ICH), University College London (UCL), 31 Guy’s and St. Thomas’ Biomedical Research Centre, 32 Guy’s and St. Thomas’ NHS Foundation Trust, 33 Hampshire Hospitals NHS Foundation Trust, 34 Health Services Laboratories, 35 Heartlands Hospital, Birmingham, 36 Hub for Biotechnology in the Built Environment, Northumbria University, 37 Hull University Teaching Hospitals NHS Trust, 38 Imperial College Healthcare NHS Trust, 39 Imperial College London, 40 Infection Care Group, St George’s University Hospitals NHS Foundation Trust, 41 Institute for Infection and Immunity, St George’s University of London, 42 Institute of Biodiversity, Animal Health & Comparative Medicine, 43 Institute of Microbiology and Infection, University of Birmingham, 44 Isle of Wight NHS Trust, 45 King’s College Hospital NHS Foundation Trust, 46 King’s College London, 47 Liverpool Clinical Laboratories, 48 Maidstone and Tunbridge Wells NHS Trust, 49 Manchester University NHS Foundation Trust, 50 Microbiology Department, Buckinghamshire Healthcare NHS Trust, 51 Microbiology, Royal Oldham Hospital, 52 MRC Biostatistics Unit, University of Cambridge, 53 MRC-University of Glasgow Centre for Virus Research, 54 Newcastle University, 55 NHS Greater Glasgow and Clyde, 56 NHS Lothian, 57 NIHR Health Protection Research Unit in HCAI and AMR, Imperial College London, 58 Norfolk and Norwich University Hospitals NHS Foundation Trust, 59 Norfolk County Council, 60 North Cumbria Integrated Care NHS Foundation Trust, 61 North Middlesex University Hospital NHS Trust, 62 North Tees and Hartlepool NHS Foundation Trust, 63 North West London Pathology, 64 Northumbria Healthcare NHS Foundation Trust, 65 Northumbria University, 66 NU-OMICS, Northumbria University, 67 Path Links, Northern Lincolnshire and Goole NHS Foundation Trust, 68 Portsmouth Hospitals University NHS Trust, 69 Public Health Agency, Northern Ireland, 70 Public Health England, 71 Public Health England, Cambridge, 72 Public Health England, Colindale, 73 Public Health Scotland, 74 Public Health Wales, 75 Quadram Institute Bioscience, 76 Queen Elizabeth Hospital, Birmingham, 77 Queen’s University Belfast, 78 Royal Brompton and Harefield Hospitals, 79 Royal Devon and Exeter NHS Foundation Trust, 80 Royal Free London NHS Foundation Trust, 81 School of Biological Sciences, University of Portsmouth, 82 School of Health Sciences, University of Southampton, 83 School of Medicine, University of Southampton, 84 School of Pharmacy & Biomedical Sciences, University of Portsmouth, 85 Sheffield Teaching Hospitals NHS Foundation Trust, 86 South Tees Hospitals NHS Foundation Trust, 87 Southwest Pathology Services, 88 Swansea University, 89 The Newcastle upon Tyne Hospitals NHS Foundation Trust, 90 The Queen Elizabeth Hospital King’s Lynn NHS Foundation Trust, 91 The Royal Marsden NHS Foundation Trust, 92 The Royal Wolverhampton NHS Trust, 93 Turnkey Laboratory, University of Birmingham, 94 University College London Division of Infection and Immunity, 95 University College London Hospital Advanced Pathogen Diagnostics Unit, 96 University College London Hospitals NHS Foundation Trust, 97 University Hospital Southampton NHS Foundation Trust, 98 University Hospitals Dorset NHS Foundation Trust, 99 University Hospitals Sussex NHS Foundation Trust, 100 University of Birmingham, 101 University of Brighton, 102 University of Cambridge, 103 University of East Anglia, 104 University of Edinburgh, 105 University of Exeter, 106 University of Kent, 107 University of Liverpool, 108 University of Oxford, 109 University of Sheffield, 110 University of Southampton, 111 University of St Andrews, 112 Viapath, Guy’s and St Thomas’ NHS Foundation Trust, and King’s College Hospital NHS Foundation Trust, 113 Virology, School of Life Sciences, Queens Medical Centre, University of Nottingham, 114 Watford General Hospital, 115 Wellcome Centre for Human Genetics, Nuffield Department of Medicine, University of Oxford, 116 Wellcome Sanger Institute, 117 West of Scotland Specialist Virology Centre, NHS Greater Glasgow and Clyde, 118 Whittington Health NHS Trust
Acknowledgments:
The authors would like to thank the participants of the DOVE study and Sister Therese McSorley and her nursing team at the NHS GG&C clinical research facility. The authors also thank Alison Hamilton, Laura Stirling and Charlie Mayor from the NHS GG&C SafeHaven team for their invaluable input in facilitating this study. We thank Paula Olmo for administrative support and Chris Robertson and Aziz Sheikh for statistical advice. The authors thank all of the researchers who have shared genome data openly via the Global Initiative on Sharing All Influenza Data (GISAID).
Acknowledgments
The authors would like to thank the participants of the DOVE study and Sister Therese McSorley and her nursing team at the NHS GG&C clinical research facility. The authors also thank Alison Hamilton, Laura Stirling and Charlie Mayor from the NHS GG&C SafeHaven team for their invaluable input in facilitating this study. We thank Paula Olmo for administrative support and Chris Robertson and Aziz Sheikh for statistical advice. The authors thank all of the researchers who have shared genome data openly via the Global Initiative on Sharing All Influenza Data (GISAID). Funding was provided by Health Data Research UK (HDR UK) for the Evaluation of Variants Affecting Deployed COVID-19 Vaccine (EVADE) study (ECT, SR, OM, CW, BJW, grant code: 2021.0155). This research is part of the Data and Connectivity National Core Study, led by Health Data Research UK in partnership with the Office for National Statistics and funded by UK Research and Innovation (grant ref MC_PC_20058). This work was also supported by The Alan Turing Institute via ‘Towards Turing 2.0’ EPSRC Grant Funding. COG-UK is supported by funding from the Medical Research Council (MRC, part of UK Research & Innovation (UKRI)), the National Institute of Health Research (NIHR) [grant code: MC_PC_19027], and Genome Research Limited, operating as the Wellcome Sanger Institute (RMB, DLR, ECT). Medical Research Council (MRC) provided funding for both the COVID-19 DeplOyed VaccinE (DOVE) study (grant code: MCUU1201412) and COG-UK (ECT). AF, JH, RO, JG, ECT, NL and DLR were funded by the Medical Research Council (MRC) (grant code: MC_UU_12014/12). W.T.H. was supported by Medical Research Council (MRC), grant codes MR/R024758/1 and MR/W005611/1. The G2P-UK National Virology Consortium was funded by UK Research and Innovation (UKRI) award MR/W005611/1 (MP, ECT, AP, DLR). D.L.R. was funded by Wellcome Trust (grant code: 220977/Z/20/Z). N.L. and B.J.W. were funded by the Biotechnology and Biological Sciences Research Council (grant codes: BBSRC, BB/R004250/1 & BB/R019843/1). J.G. was funded by a Wellcome Trust and Royal Society Sir Henry Dale Fellowship (grant code: 107653/Z/15/A).
Footnotes
↵§ Full list of consortium names and affiliations are listed in the Supplementary information.
Figures have been condensed into 4 figures and the panel 4C data on vaccine effectiveness has been updated with more recent information. The text has been modified to reduce to 4 rather than 6 figures.
References
- 1.
- 2.↵
- 3.
- 4.
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.
- 41.
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.
- 60.
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵



