
ABSTRACT
Objectives Immunocompromised patients are at risk of severe coronavirus disease 2019 and are considered a high priority for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) vaccination. Humoral vaccination response is impaired in these patients when circulating B cells are lacking. We aimed to analyze B-cell subsets at the time of vaccination to identify potential predictors of humoral vaccination response.
Methods Patients (n=120) receiving B-cell-depleting therapy (n=41), those suffering from inborn errors of immunity (n=25) and hematologic malignancies (n=56), and healthy controls (n=79) were vaccinated twice with BNT162b2 or mRNA-1273. B-cell subsets were analyzed prior to vaccination. Two independent anti-SARS-CoV-2 S immunoassays targeting the receptor-binding domain (RBD) or trimeric S protein (TSP) were performed three to four weeks after the second vaccination.
Results Seroconversion occurred in 100% of the healthy controls, in contrast to 67% (RBD) and 82% (TSP) of the patients, while only 32% (RBD) and 22% (TSP) achieved antibody levels comparable to those of healthy controls. The number of circulating naïve B cell was strongly associated with antibody levels (r=0.761, P<0.001) across all immunosuppressive treatments or conditions. In multivariable analysis, the number of naïve B cells was an independent predictor for achieving antibody levels comparable to healthy controls, and receiver operating characteristic analysis predicted that at least six naïve B cells per µL were required.
Conclusions Assessing the abundance of naïve B cells in immunocompromised patients could be useful in predicting the optimal vaccination response.
What is already known about this subject?
▸ Humoral vaccination response is impaired in patients with immunodeficiency or B-cell-depleting therapies, such as anti-CD20 antibody therapy or hematopoietic stem cell transplantation.
▸ Current recommendations advise an interval of six months between the most recent anti-CD20 antibody administration and vaccination.
What does this study add?
▸ Immunocompromised patients can achieve anti-spike protein antibody levels similar to those in healthy subjects after vaccination.
▸ Independent of concurrent immunosuppressive treatment or condition, the number of naïve B cells, but not the total number of B cells, is strongly associated with anti-spike protein antibody levels, similar to that in healthy subjects.
How might this impact on clinical practice or future developments?
▸ Assessment of the abundance of naïve B cells in immunocompromised patients could be used to schedule vaccinations for optimal antibody responses.
INTRODUCTION
Coronavirus disease 2019 (COVID-19) results in increased morbidity and mortality in immunocompromised patients.[1-3] Immunodeficiency can be primary (PID) due to underlying genetic causes such as common variable immunodeficiency or secondary (SID) resulting from hematologic malignancies (HM), immunosuppressive therapies, or hematopoietic stem cell transplantation (HSCT). In a recent study of 100 patients with COVID-19 disease, patients with PID and SID demonstrated higher morbidity and mortality than the general population, while the outcomes of individuals suffering from SID were the worst.[1] In patients with HM and COVID-19, a mortality rate of 34% [95% confidence interval (CI): 28–39] has been reported in adults.[4]
Due to the high risk of severe COVID-19 in immunocompromised patients, they are considered a high priority for COVID-19 vaccination.[5-9] However, trials of the currently approved COVID-19 vaccines have excluded individuals diagnosed with immunodeficiency or malignancy; therefore, information on the efficacy and safety of the vaccines in these patients is sparse.[10-13] Hagin et al. reported specific antibody responses in 18/26 (69%) patients with inborn errors of immunity and spike-peptide-specific T cells in 19/26 (73%) patients two weeks after administration of the second dose of BNT162b2 vaccine.[10] Braun-Moscovici et al. observed significant severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) IgG antibody levels in 227/264 patients (86%) with autoimmune rheumatic diseases (AIRD) 4–6 weeks after the second dose of BNT162b2 vaccine.[11] Other investigators confirmed these vaccination responses in AIRD patients and reported significant reductions in seropositivity rates in patients receiving B-cell-depleting therapy and glucocorticoids.[12, 13] Lack of antibody responses and significantly lower antibody levels in responders have been reported in HM patient cohorts in general[14-16] and in selected patients with multiple myeloma, chronic lymphocytic leukemia (CLL), and non-Hodgkin lymphoma.[17-21] Vaccinations have a low efficacy when administered soon after HSCT and anti-CD20 therapies.[5, 22, 23]
Recently, Mrak et al. showed that a humoral response to mRNA vaccines can also be induced in patients after therapy with rituximab as long as peripheral B cells are measurable.[24] However, the number of circulating B cells or of a certain B-cell subset associated with a humoral vaccination response comparable to healthy individuals is unknown. A marker predictive of vaccination response would aid to schedule vaccinations in the immunocompromised to achieve optimal vaccination response.
We hypothesize that specific B-cell subsets present during vaccination are associated with a humoral vaccination response. Herein, we used data from an interim analysis of the prospective, open-label, phase IV CoVVac trial (NCT04858607) to test the hypothesis.
METHODS
Study design and participants
We report the data of an interim analysis of the CoVVac trial (NCT04858607), which is an ongoing open-label, phase IV, prospective, monocentric study at the Medical University of Graz, Austria. After approval by the ethics committee of the Medical University of Graz in April 2021 (EK 1128/2021), patients with inborn errors of immunity, hematological malignancies, those receiving B-cell-depleting therapy, and healthy controls were recruited before receiving their first dose of COVID-19 vaccine. The detailed study protocol is provided in the Supplementary Information. In brief, blood was drawn before the first vaccination with BNT162b2 (BioNTech/Pfizer) or mRNA-1273 (Moderna) for peripheral blood mononuclear cell (PBMC) isolation and lymphocyte phenotyping. The second vaccination was administered 21 (BNT162b2) or 28 days (mRNA-1273) after the first. Blood sampling was performed 21–28 days after the second vaccination to analyze the COVID-19-specific antibody response as the primary endpoint.
Patient and public involvement
It was not appropriate or possible to involve patients or the public in the design, or conduct, or reporting, or dissemination plans of our research.
Lymphocyte phenotyping
Blood samples from the baseline visit were processed within 4 hours for analysis by flow cytometry. For lymphocyte phenotyping, ethylenediaminetetraacetic acid whole blood was stained for CD3, CD4, CD8, CD45, CD16, CD56, and CD19, as previously described.[25] For B-cell phenotyping, PBMCs were isolated from lithium heparin whole blood by Ficoll gradient density centrifugation. One million PBMCs were incubated with the following antibodies: CD19-VioGreen, anti-IgD-VioBlue, CD24-PerCP-Vio700, CD38-FITC, CD27-APC, CD86-PE-Vio770, CD21-APC-Vio770, and anti-IgM-PE (Miltenyi Biotec, Bergisch Gladbach, Germany). Samples were measured using a FACSLyric flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). Data were analyzed using the FACSSuite (BD Biosciences). The gating strategy is shown in Supplemental Figure 1.
Antibody assays
Serum was obtained 21–28 days after the second vaccination, aliquoted, frozen, and stored at -80°C until analysis was performed in batches. Two CE-marked serological tests were performed according to the manufacturers’ protocols to determine and quantify specific antibodies against SARS-CoV-2. A Specific IgG was determined using the Roche Elecsys anti-SARS-CoV-2 S electrochemiluminescence immunoassay targeting the receptor-binding domain of the viral spike protein using a Cobas e 801 analytical unit (Roche Diagnostics GmbH, Mannheim, Germany).[20, 24] Its quantification range lies between 0.4 and 2500 U/mL, with a cut-off of 0.8 U/mL for positivity. Specific IgG was measured by Liaison SARS-CoV-2 TrimericS IgG test on Liaison XL (DiaSorin, Saluggia, Italy), which is a chemiluminescence immunoassay quantifying antibodies that target the trimeric S protein.[21] Results are provided in binding antibody units (BAU) with a quantification range of 4.81–2080 BAU/mL. Values ≥33.8□BAU/mL were considered positive.
Statistical analysis
All statistical analyses were performed using Stata 16.1 (Stata Corp., Houston, TX, USA). Continuous data were reported as medians (25th–75th percentile) and categorical data as absolute frequencies (%). Correlations and associations between antibody response and other variables were examined using Spearman’s rank-based correlation coefficients, rank-sum test, and χ2-squared tests. R2-statistics were obtained from multiple linear regression models with antibody response as the dependent variable. The optimal cut-off to separate patients with and without vaccination response was assessed by employing a maximized Youden’s index within a receiver operating characteristic (ROC) analysis. Logistic models were used for univariate and multivariate modeling of the vaccination response. The Kruskal– Wallis test was used for continuous variables when comparing three or more treatment groups. The Kruskal– Wallis H test was used as a post-hoc test to determine between-group differences.
RESULTS
Study population
Complete data of 199 study participants who completed their full vaccination schedule were included in the efficacy analysis. Of these, 79 were healthy participants and 120 were immunocompromised patients. All study participants were vaccinated with mRNA-1273, with only two healthy individuals (1%) receiving BNT162b2. A total of 140 adverse events occurred after vaccination, with the most common being pain at the injection site, headache, fever, and fatigue. Four severe adverse events (hospitalization and death due to HM and bone fractures) were considered unrelated to vaccination. Participant characteristics are detailed in Table 1. Diagnoses, immunosuppressive treatments, and antibody responses for subgroups are listed in Supplementary Table 1.
Immunogenicity of mRNA vaccines
Antibody responses were assessed using two different assays. All healthy controls demonstrated seroconversion with high antibody titers (Roche median: 2500 U/mL; DiaSorin median: 2080 BAU/mL). In immunocompromised patients, the seroconversion rates and antibody levels were significantly lower than those in healthy controls (Table 1 and Figure 1); 67% (n = 80) and 82% (n = 98) of patients demonstrated a humoral response with antibody levels within the quantification range of Roche and DiaSorin assays, respectively. Since the clinical significance of antibody levels close to the limit of detection was unclear, we defined more stringent thresholds. In our patients, we defined a “stringent response” as reaching at least the lowest antibody level of the healthy individuals from our cohort (Roche ≥1000 U/mL; DiaSorin ≥2000 BAU/mL). According to this definition, only 32% (Roche) and 22% (DiaSorin) of patients had a stringent antibody response. This difference was not statistically significant between the two tests (P = 0.108). Patients who received anti-CD20 therapy, including the majority of patients with AIRD, had the lowest rate of stringent response (≤10%). Interestingly, patients who received HSCT demonstrated a relatively high rate of stringent response [37% (13/35)]. There was no statistically significant difference between allogeneic and autologous HSCT (40% vs. 30%, respectively; P = 0.541). The antibody levels of both assays showed an excellent correlation with each other (P = 0.915, P<0.001, R2 = 0.841; Table 2; Figure 2A). Since the Roche assay is more widely used in research and shows a good correlation with live virus neutralization tests in vaccinated individuals,[15, 20, 24, 26] we focused on the Roche assay for subsequent analyses to ensure comparability with other studies.

Immune response to SARS-CoV-2 mRNA vaccination. The scatter plot shows total Ig levels for healthy controls, immunodeficient patients and patients after anti-CD20 therapy or haematopoietic stem cell transplantation (HSCT). P is <0.001 between all groups calculated with the Kruskal-Wallis test and post hoc analysis. Lines are medians with interquartile range. The Plot was drawn with GraphPad Prism Version 9.2.0.332.
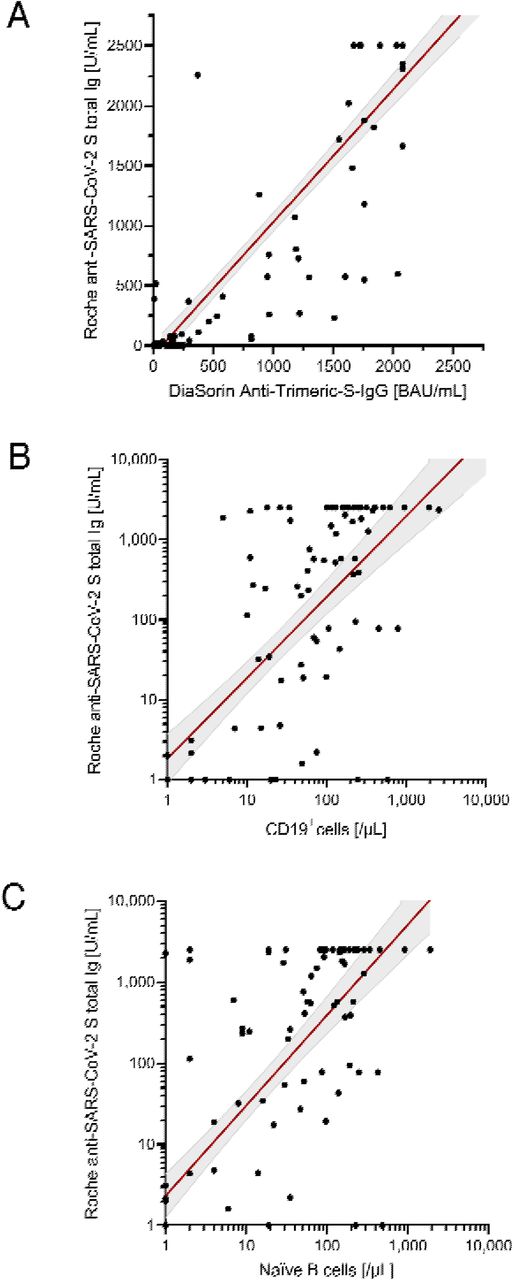
Correlation of antibody levels determined by Roche anti-SARS-CoV-2 S assay with B cells in patients. (A.) DiaSorin SARS-CoV-2 TrimericS IgG. (B.) Absolute number of B cells (CD19+ cells). (C.) Absolute number of naïve B cells. Scatter plots indicate a linear regression line including a 95% confidence interval. In case of A. and B., regression line corresponds to transformed data using x=log((x+1) and y=log((y+1)), respectively.
Correlation of antibody levels with the amount of B-cell subsets
The total number of B cells and all B-cell subsets were positively correlated with the antibody levels in all patients (Roche: P = 0.739, R2 = 0.001, P<0.001; Table 2; Figure 2B). In this analysis, the absolute number of naïve B cells showed the highest correlation with antibody titers (Roche: P = 0.761, R2 = 0.153; Figure 2C). Furthermore, this analysis also indicated that the time between the last B-cell-depleting therapy and vaccination was also a significant factor correlating with antibody levels. Extending this correlation analysis to the entire study population (Supplementary Table 2; Supplementary Figure 2), the influence of naïve B cells remained highly significant (Roche: P = 0.636, P<0.001, R2 = 0.123). However, because the age of healthy subjects was significantly lower than that of patients, age appeared to be a significant factor negatively correlated with anti-SARS-CoV-2 antibodies after vaccination in the entire study population (Roche: p = −0.209, R2 = 0.057).
To support the results of our correlation analyses, we established a model of stringent vaccination response prediction for the entire study population based on the results of the Roche assay using the area under the ROC curve (AUC; Supplementary Table 3). This model suggests that the total number of B cells (AUC: 0.87, 95% CI: 0.79–0.92), naïve B cells (AUC: 0.82, 95% CI: 0.73–0.88), pre-switch memory B cells (AUC: 0.84, 95% CI: 0.76–0.89), and switched memory B cells (AUC: 0.85, 95% CI: 0.79–0.91) can best describe the response to vaccination in the entire study population, and that age does not play a significant role.
Naïve B cells show the strongest association with stringent response
To test the association of variables with vaccination response determined by the Roche assay, we performed univariate and multivariate linear regression analyses on the entire study population using the same variables from the correlation analyses.
In univariate analysis (Table 3), the variables found to be significantly associated with any antibody response in patients included interval in days from the last B-cell-depleting therapy, naïve B-cell count, pre-switch memory B-cell count, and absolute number of plasmablasts. The same correlation was also found for stringent antibody responses, except for the correlation with pre-switch memory B cells. The absolute number of these B-cell subsets was significantly lower in patients than in healthy controls, in patients after monoclonal anti-CD20 antibody therapy compared to HSCT, and in patients without seroconversion compared to those with any antibody response (not shown) or stringent response (Supplementary Figure 3). The variables of immunodeficient patients stratified by vaccination response are shown in Supplementary Table 4. In the univariate analysis of the entire cohort (Supplementary Table 5), the same B-cell subsets remained significantly associated with vaccine response. In addition, we found a negative correlation with age, but only for the stringent antibody response [odds ratio (OR): 0.69 per 10-year increase, 95% CI: 0.54–0.87, P = 0.001].

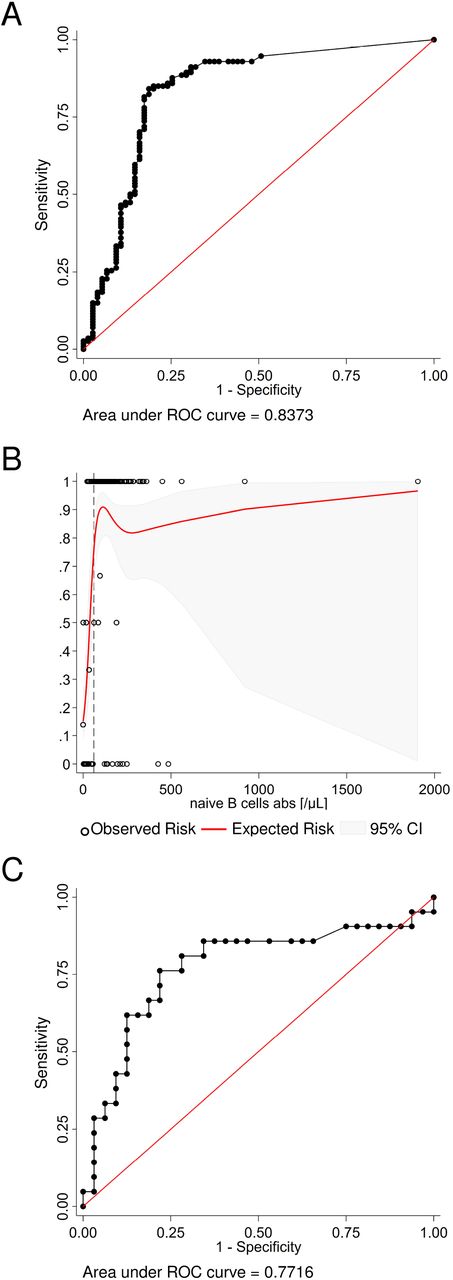
Exploratory analyses estimating the number of naïve B cells and the interval to the last B-cell depleting therapy required for a vaccination response. (A.) The receiver operating characteristic (ROC) analysis curve for naïve B cells differentiating stringent antibody response vs no or any vaccination response in the whole study population (n=189) shows excellent discrimination. (B.) A non-linear risk model based on the observed risk of seroconversion was created to estimate the minimum number of naïve B cells required for a stringent vaccination response. Independently, the best discriminatory cut-off (dashed line) was determined with the Youden’s index from the ROC curve. Both models predict that six or more naïve B cell per µL are required to generate a stringent vaccine response. (C.) The ROC analysis curve for the interval since the last B-cell depleting therapy up to 365 days (n=53) differentiating any seroconversion vs no vaccination response. The optimal cut-off is an interval of 116 days or more.
In multivariable analysis for stringent response, only the number of naïve B cells was an independent predictor (OR: 1.07 per 10-µL increase, 95% CI: 1.02–1.12, P = 0.009). The only independent predictor of any seroconversion was plasmablast count (OR: 4.42 per 1-µL increase, 95% CI: 1.30– 15.01, P = 0.017). The interval since the last B-cell-depleting therapy had to be excluded because this variable was not applicable to all patients.
The multivariable analyses for the entire study population are shown in Supplementary Tables 6 and 7. The absolute number of naïve B cells remained an independent predictor of stringent antibody response (OR: 1.14 per 10-µL increase, 95% CI: 1.08–1.20, P<0.001), while age was not associated with vaccination response.
Exploratory analyses
As naïve B cells were the only B-cell subset independently associated with stringent antibody response, we were interested to determine whether our dataset allowed us to estimate a naïve B cell count threshold for stringent antibody response. ROC analysis and non-linear risk modeling predicted that at least six or more naïve B cells per µL were required to generate a stringent vaccine response (Figures 3A, B).
A second exploratory ROC analysis was undertaken to determine the minimum interval between the last B-cell-depleting therapy, i.e., anti-CD20 antibody therapy or HSCT up to 365 days (n = 53), and vaccination differentiating any seroconversion versus no vaccination response. The optimal cut-off in this population was 116 days or more (Figure 3C).
DISCUSSION
We investigated the relationship between B-cell subsets and humoral vaccination response to COVID-19 mRNA vaccination in immunocompromised patients and healthy controls. Recent publications show diverging antibody levels depending on the underlying diseases of the investigated population, age, and immunosuppressive treatments. Our findings confirm those from recent literature, which demonstrated that immunocompromised individuals show significantly lower and more heterogeneous antibody titers than healthy controls.[10, 11, 15, 17-21, 24]
In one of the largest studies published on COVID-19 vaccination in patients with HM, Maneikis et al. reported lower median anti-S1 IgG responses after two BNT162b2 vaccine doses in 653 patients with HM than 69 healthy healthcare workers.[16] However, the response was highly heterogeneous, with many patients reaching an antibody response similar to that of healthy controls. Even in the group treated with anti-CD20 therapy, 19% had anti-S1 IgG antibodies in the range of healthy controls. While the factors responsible for such a favorable humoral vaccination response are unknown, a similar heterogeneity in vaccination response has been reported for PID,[10] AIRD,[12] and anti-CD20 therapy in several studies.[15, 16, 20, 27] Recently, Mrak et al. found that the presence of circulating B cells was essential for the humoral response to BNT162b2 after rituximab therapy.[24] While they found a moderate correlation with the relative abundance of circulating B cells, they state that the mere presence of peripheral B cells allows seroconversion irrespective of the B cell count.[24] Interestingly, in our univariate analysis, the absolute number of CD19+ B cells prior to vaccination was not associated with anti-SARS-CoV-2 response in patients, whereas naïve B cells, pre-switch memory B cells, and plasmablasts were associated with seroconversion. However, only naïve B cells were independent predictors of stringent vaccination responses (i.e., similar to healthy subjects) in the multivariable analysis.
The abundance of circulating naïve B cells predicted stringent vaccination responses in patients across a wide range of diseases and treatments, suggesting the functional importance of these cells in the humoral immune response. Indeed, the production of specific antibodies to a novel antigen relies on the presence of antigen-specific B cells within the naïve B cell pool.[28] Thus, a drastically contracted pool of naïve B cells reduces the chance of harboring B cells with a B-cell receptor of high avidity to the antigen. Only B cells with a high B-cell receptor/antigen avidity can interact with T follicular helper cells successfully and subsequently undergo somatic hypermutation to develop an optimal antibody response.[29] A lack of these cells results in low antibody titers of poor quality. Therefore, the association of the humoral vaccination response with the abundance of naïve B cells is most likely a causal relationship.
Establishment of protective humoral immunity following vaccination may be impaired until the naïve B cells repopulate.[5] Therefore, current recommendations suggest time intervals of up to six months between the most recent rituximab administration and vaccination.[30-32] Our data confirm that time intervals play a role in seroconversion, with an estimated cut-off of 116 days from the last B-cell-depleting therapy and vaccination. Nevertheless, several independent statistical models suggest that B-cell subsets, the targets of these therapies, are the actual predictors of vaccination response. In a larger cohort, Mrak et al. found that the time since the last rituximab treatment correlated with B-cell frequency but was not an independent predictor of seroconversion.[24]
Our study had several limitations. These include single-center design and limited representation of some patient cohorts that do not allow clear conclusions on seroconversion rates among less common entities or less frequently used treatment strategies. Moreover, we cannot make any statements about the persistence of the vaccination response at this point. Our study relies on the measurement of antibodies as a surrogate for immunity to COVID-19. However, our results do not significantly differ between the two internationally deployed anti-spike protein serological assays for detecting either total IgG or IgG. Both tests showed a high correlation with surrogate neutralization tests, and Roche’s assay correlated well with live virus neutralization tests in vaccinated individuals.[24, 26] The strength of our study is its prospective design and the introduction of the stringent vaccination response as a potentially more relevant concept than seroconversion.
In summary, the humoral response to the SARS-CoV-2 mRNA vaccine is impaired in immunocompromised patients. The abundance of circulating naïve B cells is strongly associated with a normal antibody vaccine response across different conditions and therapies. Therefore, measuring naïve B cells may predict the humoral response to COVID-19 vaccination. Further research is needed to confirm these findings vaccinating immunocompromised individuals against COVID-19 and other pathogens.
Data Availability
All data will be shared upon request.
Author contributions
IH, ES, PS, SH, HG, IS, and MHS designed the study. ES, IH, PF, NS, JF, BD, BU, AG, MM, CU, MK, CE and BK collected clinical samples and/or data. PF and BK performed antibody assays. ES and SH analyzed the data and performed the statistical analysis. ES, SH, IH, PS, HG, IS, and MHS critically reviewed and discussed the results. ES, IH, PF, HS, HG, and MHS wrote the first draft. All authors reviewed the draft and approved the final version of the manuscript.
Competing interests
None of the contributing authors have a conflict of interest, including specific financial interests, relationships, and affiliations relevant to the topic or materials covered in the manuscript.
Funding
None
Acknowledgments
We thank Pia-Carina Gallaun and Julia Lodron for technical assistance. The samples/data used for this project were processed with the help of the Biobank Graz of the Medical University of Graz, Austria.




{kind=link}
{kind=link}
{kind=link}