
Abstract
Fragile X Syndrome (FXS) is a monogenetic form of intellectual disability and autism in which well-established knockout (KO) animal models point to neuronal hyperexcitability and abnormal gamma-frequency physiology as a basis for key illness features. Translating these findings into patients may identify tractable treatment targets. Using a minimum norm estimate of resting state electroencephalography data, we report novel findings in FXS including: 1) increases in gamma activity across functional networks, 2) pervasive changes of theta/alpha activity, indicative of disrupted thalamocortical modulation coupled with elevated gamma power, 3) stepwise moderation of these abnormalities based on female sex and genetic mosaicism, and 4) relationship of this physiology to intellectual disability and anxiety. Our observations extend findings in Fmr1-/- KO mice to patients with FXS and raise a key role for disrupted thalamocortical modulation in local hyperexcitability, a mechanism that has received limited preclinical attention, but has significant implications for understanding fundamental disease mechanisms.
Introduction
During spontaneous brain recordings, measured fluctuations in neural activity reveal highly structured functional dynamics in resting-state networks (RSN)1 that can be characterized with millisecond resolution using dense-array electroencephalography2. In neurodevelopmental disorders (NDDs), higher order cognition, emotional regulation, and sensorimotor functions are notably altered3. However, the cortical activity that engender these changes remain largely unknown. In monogenetic NDDs, where animal models are well established, slice and unit electrophysiology studies of Fmr1-/- knockout mice (KO) have led to interest in in vivo dense array electroencephalography (EEG), paralleling similar interest of neural dysfunction in clinical research. Thus, translational electrophysiological investigations now provide a novel strategy for developing targets for clinical intervention and trans-species biomarkers for predicting and tracking drug effects4.
Fragile X Syndrome (FXS) is a promising target as clinical and preclinical research suggests neural hyperexcitability as a key illness feature5, 6. FXS involves reduced production of fragile X mental retardation protein (FMRP) that is encoded by FMR1 on the X chromosome. FMRP is an RNA binding protein that regulates protein synthesis at synapses, maintains synaptic plasticity, and is crucial for higher order cognitive functions7. Reduced levels of FMRP result in near universal intellectual disability with a high prevalence of anxiety disorders, communication impairments, sensory hypersensitivities, and autism. Fmr1-/- KO display evidence of heightened cortical excitability and altered excitatory/inhibitory (E-I) homeostatic mechanisms8, 9. For example, Fmr1-/- KO have increased susceptibility to audiogenic seizures and enhanced gamma oscillatory activity (> 30 Hz) as measured by murine multielectrode array10-12. Microcircuit experiments in Fmr1-/- KO cortical slices have established excessive spontaneous network spiking, enhanced gamma power, and altered gamma synchrony between cortical L2/3 and L4 cortical layers8, 13.
EEG from Fmr1-/- KO and patients with FXS have increased spontaneous, broadband gamma power and diminished narrow-band gamma response to auditory stimuli11. These preliminary investigations also indicate that excess intrinsic gamma activity is associated with electrophysiological alterations during sensory evoked14 and speech production tasks15. However, human EEG studies of FXS have been limited by small samples (n<25), under representation of females, failure to ascertain mosaic status in males, and lack of source and functional network modeling16. Gamma oscillations hold special interest for NDDs because of their relation to cortical excitability and cognition17. While studies of gamma power face challenges using scalp recordings, the reconstruction of source activations from scalp EEG signals in concert with independent component analysis (ICA) can effectively improve spatiotemporal resolution by reducing volume conduction effects and contamination of high-frequency activity by scalp musculature18.
Here, in a relatively large sample of FXS patients, we report on neural activity at the cortical source level and the relationships of this activity with disease features. We demonstrate regional and network-based broadband gamma elevations related to severity of intellectual disability and neuropsychiatric symptoms. We also demonstrate a marked attenuation of alpha activity (with a concomitant increase in theta activity) and altered coherence of this low-frequency activity with gamma activity, which is highly suggestive of disruptions of thalamocortical modulation of neocortical hyperexcitability. Comparisons of full-mutation male and female FXS participants (obligate mosaics) indicated that modest FMRP expression in females appear to mitigate excessive broadband gamma activity but certain alterations in thalamocortical function persist. The present findings provide a comprehensive profile of resting-state cortical activity in a monogenetic NDD that confirms and extends mechanistic gamma findings of the FMR-/- KO at the local circuit level, novel insights into low-frequency thalamocortical activity in influencing neocortical hyperexcitability, and strong evidence of the relation of these alterations to core clinical features of FXS.
Results
Source-estimation reveals broadband spectral alterations
Eighty-seconds of artifact-free data were analysed for each participant based on blinded, semi-autonomous pre-processing (see Methods and Figure 1a) from five-minutes of continuous EEG recording from 141 study participants: 70 individuals with a genetic diagnosis of FXS (without seizures or on antiepileptics) and 71 age- and sex-matched typically developing control participants (see Table 1 and Table S1). Non-verbal intelligence quotients (NVIQ) were severely reduced in males with FXS (M=22.1, 95% CI [12-32.2]) and to a lesser degree in females (M=59.2, 95% CI [47.1-71.3]; see Figure 1b).
FSIQ, Full Scale IQ; NVIQ, Non-verbal intelligence quotient; VIQ, verbal intelligence scale; SCQ, Social Communication Questionnaire; WJ-3, Woodcock III Tests of Cognitive Abilities; adj. p, p-value following Bonferroni correction.

Spectral alterations suggestive of neocortical hyperexcitability in FXS. Scalp and source-localization of dense array electroencephalography (EEG) suggest broadband power disruptions in frequency oscillations in FXS (n=70) compared to age- and sex-matched controls (n=71). (A) Principal steps of the investigative method: Following blinded preprocessing, artifact-free EEG data and a cortical lead field matrix was used to construct a weighted minimum norm estimate (MNE) to reconstruct source estimations. Current source density (CSD) by frequency band was used to calculate 1) vertex by vertex differences between groups, 2) generate contrast independent time series for Desikan-Killany atlas nodes based on the atlas, and 3) hierarchical groupings into EEG-defined resting-state networks (RSN). (B) Subject-level scatter plot of intellectual disability as estimated by non-verbal intelligence z-scores (NVIQ) by allele group. (C) Frequency spectrogram of mean scalp EEG relative power (thick lines) with 95% confidence intervals (dotted tracings) by group and sex. Peak frequency is reduced in FXS males (M=8.0 Hz, SD=1.8) and females (M=8.6 Hz, SD=1.7) compared to control males (M=9.3 Hz, SD=1.8) and females (M=9.6 Hz, SD=1.7) with a prominent effect of Group: F1,133 = 38.24, p = 1.5×10−9. No interaction effect or effect of age (F1,133 =.02, p=.89) on peak alpha frequency was evident. Inset: Isolated depiction of gamma frequencies (30-90Hz) illustrating gross elevation of gamma activity in FXS males. (D) Group level t-maps depicting FXS – control, vertex-by-vertex relative power differences by frequency band superimposed on brain surface models. Warmer (FXS > Control) and cooler (FXS < Control) color scale represents significant t-values (non-significant values as gray). (E) Boxplots displaying median and interquartile range of relative power by frequency band (split into subplots by frequency band to optimize scale), averaged for each significant ROI from group level t-maps. Subject level data is superimposed as a scatterplot.
Comparisons of topographic relative power from scalp EEG (see Figure S1) and peak alpha frequency (see Figure 1c) are suggestive of cortical hyperexcitability (high broadband gamma) and reduced long-range modulation (reduced alpha peak frequency) in FXS. The source distributions of these alterations have remained ambiguous. To estimate the distribution of neural currents, we constructed a depth-weighted minimum norm estimate (MNE) model in Brainstorm19. Scalp electrodes were co-registered to an averaged Montreal Neurologic Institute (MNI) ICBM152 common brain template (see Figure 1a). Current source density (CSD) was computed across a resultant triangular mesh (15,002 vertices) which represented the cortical envelope. The Desikan-Killiany (DK) atlas20 was used to parcellate vertices into 68 cortical nodes (see Table S14).
Vertex-level comparison of FXS and control groups (see Figure 1d) indicated that FXS participants demonstrated widespread cortical elevation of delta (2-3.5 Hz), theta (4-7.5 Hz), gamma1 (30-55 Hz), and gamma2 (65-90 Hz) activity. Significant increases in gamma activity were noted across bilateral temporal lobes with limited involvement of bilateral parietal and occipital lobes. Alpha1 (8-10 Hz) and alpha2 (10.5-12.5 Hz) activity was decreased bilaterally throughout the neocortex, with relative sparing noted in frontal cortex. Beta (13-30 Hz) activity was attenuated across frontal and posterior regions, with elevated activity in R temporal lobe.
Clinical Correlations: In FXS, age-adjusted correlations were identified between gamma1 power and NVIQ (rho= -.27, p<.05, n=64), and alpha2 power and social communication (rho= .33, p<.01, n=62), obsessive compulsive symptoms (rho= -.43, p<.001, n=64), irritability (rho= -.28, p<.05, n=61), and hyperactivity (rho= -.30, p<.05, n=61) (see Table S2).
Group comparisons of cortical parcellation maps
Next, using a linear mixed effect model (LME) we identified group differences in log relative power at the atlas node level (F402,66025=1.14, p=.03; see Methods). Corrected contrasts between groups were tabulated including centroid coordinates and statistical significance (see Table S3) and top significant contrasts summarized in Table 2.
Estimates (Est.) reflect FXS-control with values greater than one indicating FXS greater than control. FDR, false discovery rate; L, Left; R, Right; C, Central; L, Lingula; P, Parietal; T, Temporal; t, t-value; PF, prefrontal; MNI, Montreal Neurological Institute X, Y, Z coordinates of ROI centroid; STS, superior temporal sulcus.
Within the theta band, 29/68 nodes survived correction with the largest increases in FXS compared to controls being found within bilateral paracentral and posterior cingulate regions, and in right (R) precuneus. For the alpha1 band, temporal regions had reduced power in FXS including the bilateral temporal pole and entorhinal cortex. Alpha2 power was globally reduced (all 68 nodes) in FXS with the largest reduction seen in the bilateral paracentral and posterior cingulate regions. Fifteen out of 68 nodes demonstrated a reduction within the beta band in FXS including five nodes in across the prefrontal cortex region. Gamma1 (15/68 significant nodes) and gamma2 (13/68 significant nodes) power was increased in FXS. The most robust elevations were seen in bilateral superior temporal sulcus and transverse temporal gyri, and R isthmus of the cingulate gyrus.
Evidence for thalamocortical abnormalities in FXS
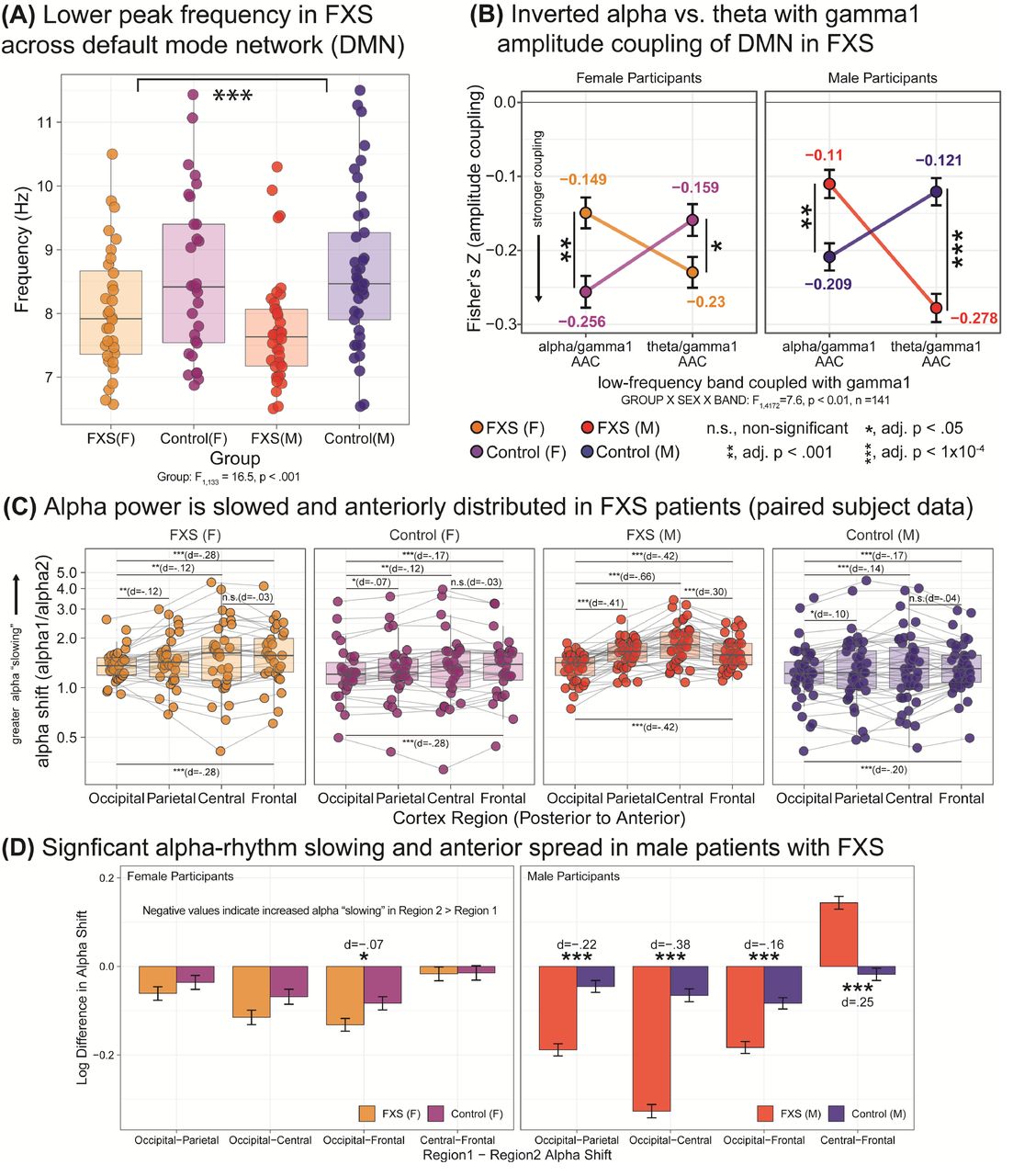
Cortical alpha rhythms in neocortex are primarily generated by thalamic input with some contributions from other neocortical sources. A constellation of findings known as thalamocortical dysrhythmia (TCD) has been observed in states such as epilepsy, tinnitus, and severe neuropsychiatric disease and supported by direct recordings from human thalamus and animal models21, 22. EEG evidence of suggestive of TCD is present in FXS including consists of 1) leftward shift of peak alpha frequency towards the theta frequency, 2) centralized distribution (known as anteriorization) of “slow” alpha frequencies, 3) increased spontaneous gamma activity, and 4) inversion of gamma to theta and alpha power coupling relationships. Reduced peak frequency in FXS: Similar to scalp EEG, the average alpha peak frequency computed from DMN nodes was reduced in in FXS (see Figure 2a; FXS males, M=7.7 Hz, SD=.88; FXS females, M=8.1 Hz, SD=.96) compared to control groups (Control males, M=8.7, SD=1.2 Hz; Control females, M=8.6, SD=1.3 Hz; F1,133= 16.5, p<.001).

Features supporting thalamocortical dysrhythmia (TCD) in FXS patients. In several neuropsychiatric conditions, pathological slowing of alpha oscillations is observed across M/EEG and intraoperative LFP recordings suggesting abnormalities in thalamocortical activity. Fmr1-/- KO studies demonstrate dysregulated activity at the thalamic level leading to aberrant entrainment of cortical networks, but thalamocortical signaling is poorly understood in FXS patients. (A) Boxplot (median and interquartile range with superimposed subject-level data) demonstrating leftward shift of mean peak alpha frequency within DMN nodes in FXS patients compared to age- and sex-matched controls. (B) Interaction plot of theta and total alpha (8-13.5 Hz) cross-frequency amplitude/amplitude coupling (AAC) with gamma1 power demonstrating that amplitude/amplitude coupling relationships are inverted in patients versus controls (vertical black bars, significant following FDR-adjusted post-hoc testing). (C) Slowing of alpha power in males with FXS with prominent “anteriorized” distribution. Bar plots (median and interquartile range with superimposed subject-level data) depict group statistical comparisons of alpha1/alpha2 ratio (higher values indicate greater proportion of low-alpha power) by group and sex across midline cortical regions (horizontal black bar, post-hoc testing: *, adj. p < .05; ***, adj. p. < 1×10−5). (D) Bar plots (mean ± standard error of least-squared mean estimates) depicting between group contrasts demonstrate males with FXS have greater proportion of “slow” alpha activity in central and frontal cortices than age-matched controls (horizontal black bars, FDR-adjusted, post-hoc testing between FXS versus matched control groups). FDR-adjusted p values, *, adj. p < .05; ***, adj. p. < 1×10−5.
Predominance of Theta-Gamma coherence in FXS
Cross-frequency coupling (CFC) examines the association between the magnitude of the recorded signal over time and between frequency bands23. Higher coherence between theta and gamma scalp EEG activity compared to alpha-gamma activity has been detected in FXS, while the reverse was seen in control participants24. This inversion has been associated with disruptions in task-related functional activity and cognitive processing25. We assessed mean CFC (expressed in normalized Fisher’s Z-based correlation coefficients) between all pairwise nodes within the DMN network. A significant inversion of alpha/theta AAC relationships (F1,4172=7.6, p<.01) favouring theta-gamma over alpha-gamma coupling was found in FXS (see Figure 2b).
“Anteriorized” Distribution of Alpha Slowing in FXS
To further clarify the distribution of the “slowed” alpha power, we calculated alpha-shift as the ratio of alpha1 to alpha2 relative power, with a higher ratio indicating greater alpha1 compared to higher alpha226 (see Table S5). We observed a significant third order interaction effect (F3,4359=17.6, p<2.5×10−11; no effect of age F1,136=.84, p= .35) indicating that males with FXS had a greater alpha shift in central nodes compared to occipital, parietal, or frontal nodes (Cohen’s d=.3-.7; see Figure 2c-d and Table S6,7).
Oscillatory activity in cognitive and affective networks
Small sample functional magnetic resonance imaging (fMRI) studies have demonstrated reduced connectivity across brain networks in FXS27. Dynamic EEG RSNs defined by connectivity analysis and within EEG source space allows for study at much higher temporal resolution, allowing higher frequencies of brain activity not accessible by fMRI28. We reconstructed five RSNs: default mode network (DMN), default attention network (DAN), salience affective network (SAN), visual network (VIS), and the auditory network (AUD; see Supplement Table 13). Prominent group differences (FXS-Control) in RSN activity by frequency band emerged at the node level in DMN (F24,43151=3.06, p<1×10−4) and in other networks. Bar plots summarizing RSN comparisons in males with FXS (with control males and affected females) are found Figure 3b. Between and within group comparisons of females with FXS and female controls are in Figure S5). Age-adjusted, FDR-corrected clinical correlations were calculated based on mean RSN and node level activity for low (theta, alpha1, and alpha2) and high frequency bands (beta, gamma1, and gamma2).

Hyperexcitability (reduced alpha and increased gamma power) in default mode network (DMN) and relationship to intellectual disability in FXS. (A) FDR-corrected group-level t-maps depicting significant differences in node-by-node relative power across DMN. Size of node is scaled by absolute t-value to enhance visualization of contrasts. Gray nodes indicate non-significant contrasts. (B) Profile of resting state network (RSNs) differences of males with FXS compared to control males and females with FXS. Obligate mosaicism present Fmr1 in females with full mutation FXS may attenuate elevated gamma power. Group-level contrasts in log-normalized relative power by frequency band of functional resting-state networks (RSNs) depicted in bar plots of least-squared mean estimates (mean ± standard error). Black outlined bars indicate frequency band and network pair is statistically significant (p < .05, FDR corrected). Gray outlined bars did not reach statistical significance. (C) Non-verbal intelligence quotient (NVIQ) is a general measure of intelligence which is not dependent on verbal abilities. NVIQ is inversely related to mean gamma1 power of DMN nodes across all FXS subjects, with the association primarily driven by full mutation males with FXS. (D) Severity of anxiety is inversely associated with alpha2 power derived from the visual network. Resting state network abbreviations: DMN, default mode network; DAN, dorsal attention network; SAN, salient affective network; VIS, visual attention network; AUD, auditory network.
Cognitive Networks (see Figure 3a for DMN and Figure S3 for DAN and SAN networks)
Default Mode Network (DMN): Functional and structural alterations of DMN have been associated with intellectual deficits in FXS27. Group differences: Theta band: elevated in 7 of 14 nodes in FXS including aspects of the cingulate cortex (R anterior, p<.05, bilateral posterior, p<.01, bilateral isthmus (iCC), p<.05), and bilateral precuneus, p<.05. Alpha: Alpha1 was decreased in 11 of 14 nodes (p<1×10−5). Alpha2 was decreased across all 14 nodes, p<1×10−5. Gamma: Gamma1 elevated in 4 of 14 nodes (bilateral parahippocampus, p<.05, and bilateral isthmus of the cingulate cortex, p<.05) and gamma2 elevated in 2 of 14 nodes (R parahippocampus, p<.05, and R iCC, p<.05).
Dorsal attention network (DAN): Attentional abilities are impaired and prognostic in FXS29. DAN regions are associated with top-down regulation of task activity or sensory processing. Group differences: Theta: Elevated in two out of 11 nodes in FXS (L pars opercularis, p<.05 and L anterior cingulate, p<.05). Alpha: Alpha1 power was reduced in 5/11 nodes, p<.001, and alpha2 was reduced across all nodes, p<1×10−5. Gamma: Gamma1 and gamma2 power was similarly elevated in 2 of 11 nodes (p<.05) including L middle and inferior temporal gyrus.
Salience Affective Network (SAN): Regions within the SAN are involved in the integration of sensory, emotional, and cognitive information including interoception and subjective awareness30. Theta: Elevated in 5 of 8 nodes in FXS (L insula, p<.05, bilateral supramarginal gyrus (SMAR), p<.05, and bilateral middle frontal gyrus, p<.05). Alpha: Alpha1 power is decreased in 3 of 8 nodes (bilateral insula, p<.05, and R SMAR, p<.05) and alpha2 power was significantly decreased across all nodes (p<.001). Gamma: Two out of 11 nodes had elevation of gamma power in FXS including the bilateral SMAR (gamma1: bilateral, p<.01; gamma2: R only, p<.01).
Clinical correlations of cognitive networks
Average DMN gamma1 power was inversely correlated with NVIQ in all FXS participants (rho=-.32, p= .01, n=64) primarily in full mutation, non-mosaic males (FM) (rho=-.47, p=.028, n=22), but not in affected males with mosaicism (MM) (rho=-.0091, p=.99, n=11) or affected full mutation females (FF) (rho=-.15, p=.41, n=31) (see Figure 3c). Significant node-by-node correlations of DMN, DAN, and SAN RSNs are presented in Supplemental Tables 9-11. In the FM group, gamma1 power of the L parahippocampus (DMN) had the strongest correlation with NVIQ (rho=-.63, p=.002). Severity of anxiety and obsessive-compulsive symptoms across all FXS subjects and FM were significantly inversely associated with alpha1 and alpha2 power including in the orbitofrontal gyrus, bilateral parahippocampus, and temporal regions.
Sensory Networks (see Figure S3)
Consistent with polymodal sensory abnormalities in FXS, slice preparations from the auditory and visual cortices in Fmr1-/- KO demonstrate immature dendritic spines, reduced synaptic pruning, circuit level hyperexcitability and altered neural synchrony31.
Auditory (AUD): Group Differences: Theta: No difference. Alpha: Both AUD nodes including the R and L superior temporal gyrus (STG) demonstrated decreased alpha1, p <.05, and decreased alpha2 power (R STG, p<.001 and L STG, p<.05). Gamma: Increased gamma1 (L STG only, p<.05) and gamma2 power (R STG, p<.05 and L STG, p<.05).
Visual (VIS): Group differences: Theta: Elevated in 2 out of 8 nodes (bilateral cuneus, p<.05) Alpha: Alpha1 was reduced similarly in 6 of 8 nodes, including bilateral fusiform, lateral occipital gyrus, and lingual gyrus, p<.05. Alpha2 was decreased in 8/8 nodes. Gamma: gamma1 was elevated in the R fusiform only, p<.05.
Clinical Correlations of sensory networks
Mean gamma1 power in AUD (ALL: rho=-.35, p<.01, n=64; FM only: rho=-.49, p<.05, n=22) and VIS (ALL: rho=-.43, p<.001, n=64; FM only: rho=-.63, p<.01, n=22) were inversely correlated with NVIQ. Mean alpha2 power in VIS but not AUD was inversely associated with ADAMS anxiety score in full mutation males (rho= -.55, p<.01, n=27; see Figure 3d). Corrected sensory RSN nodal clinical correlations are presented in Table S12 and 13. Multiple nodes within the VIS and AUD demonstrated beta and gamma power correlations with NVIQ across all FXS subjects and the FM subset. Alpha power of nodes within the visual, but not auditory RSN, was correlated with anxiety and OCD symptoms. In the FM group, the L fusiform gyri demonstrated the strongest association with anxiety severity (rho=-.63, p < .05, n=27) and OCD symptoms (rho=-.59, p<.05, n=27).
Intermediate Electrophysiological Alterations in Females with FXS
We conducted an LME which compared log relative power of five cortical regions (see Table S14) by frequency band. A 4th-order interaction effect of sex x group x region x frequency band was identified, F24, 66839=3.94, p<1×10−4 (see Figure S4 and Table S4). Compared to affected females, males with FXS had similar theta power changes but greater increases in gamma1 and gamma2 power across all cortical regions, as well as reduced alpha1 (frontal and temporal) and alpha2 (central, frontal, and temporal) activity (see Table 3). Thus, females with FXS had lower local hyperexcitability (gamma power) and more typical patterns of thalamocortical influence as indexed by alpha activity32. Sex Differences in RSNs: We further studied the effect of group and sex on RSN activity identifying a 4th-order interaction Group x Sex x RSN x Frequency (F24,43151=3.06, p<1×10−4). Figure 3b depicts differences between males with FXS and females with FXS (for other group contrasts see Figure S5). Females with FXS had similar alpha2 levels in sensory RSNs, but had increased alpha2 power in the DMN, DAN, and SAN. Gamma1 and gamma2 power was elevated in all RSN in males with FXS compared to females with FXS.
Total number of significant regions out of total regions (central, frontal, occipital, parietal, and temporal) denoted in parentheses. FXS, Fragile X Syndrome; TDC, Typically Developing Controls.
Discussion
Despite achievement of significant advances in mechanistic understanding of monogenic NDDs in animal models, translation of findings to patient-oriented research and successful treatment development have lagged. Development of translational biomarkers is needed to bridge this divide and spur progress in drug development4. In the present study, using source estimation of dense EEG resting-state data and neural network modeling in FXS, in a sample much larger than previous studies, yielded three novel findings. First, we observed clinically associated increases in gamma activity from cortical sources that varied across functional networks. Next, in the first study including a significant number of females with FXS, we identified a pattern of similarities and areas of reduced abnormalities relative to males. Third, we observed marked disruptions of theta/alpha activity indicative of thalamocortical dysfunction, which was coupled with elevated gamma power. Some observations, such as excessive gamma activity confirm findings in Fmr1-/- KO, but long-range coupling disturbances suggests an important role for altered thalamocortical modulation of local circuit excitability in the neocortex. This systems-level feature has received limited attention in preclinical research and may be an important mechanism for neocortical hyperexcitability.
In the present study, after blinded pre-processing to minimize myogenic artifact, elevations in gamma power were evident across bilateral temporal lobes and portions of the parietal and occipital lobes relative to sex- and age-matched controls. Node-by-node comparisons identified increased relative gamma power within superior temporal sulcus, fusiform region, cingulate cortex, supramarginal gyrus, and parahippocampus in FXS. Elevated resting gamma activity may disrupt circuits by decreasing signal-to-noise ratio therefore information processing.
We additionally observed widespread alterations in spectral power across RSNs that are relevant to cognition and brain disorders. We primarily focused on the DMN as it displays the highest level of fractional occupancy during resting EEG and serves as a key transition between other task-active RSNs33. Within the DMN, elevated gamma power was observed in the parahippocampal and cingulate gyri. These regions are recognized as among the strongest “hubs” or connector nodes as determined by the highest number of connections and the degree of network collapse when the node is removed2, 34. Interestingly, these regions also demonstrated the highest degree of correlation between elevated gamma power and intellectual disability within full mutation males.
Sensory networks are of particular interest, as these have been intensively studied in the Fmr1-/- KO mouse model. Auditory and visual network nodes in FXS males displayed elevated intrinsic broadband gamma power, consistent with findings in the mouse model. This could contribute to altered sensory processing, as seen in our prior observations of a reduced ability of FXS patients to mount a 40 Hz phase-locking response to auditory stimuli, which was related to excess background gamma power14. Additionally, we have found less intelligible speech related to increased broadband phasic gamma power immediately prior to talking in frontal language cortex in FXS patients15. These findings point to the functional significance of excessive phasic and intrinsic broadband high-frequency activity in FXS. In the visual cortex, narrow-band gamma oscillations support features processing such as size, contrast, and spatiotemporal aspects35. Reduced FMRP expression has been associated with impairments in contrast sensitivity at low spatial/high temporal frequency and motion coherence, implicating disruption of magnocellular (M) pathway36. Mechanistically, increases in broadband gamma in cortex coupled with reduced thalamocortical modulation (as discussed below) would be expected to increased variability of spike timing and reduced spatiotemporal selectivity in sensory cortices with adverse functional effects on sensory processing.
The translational nature of electrophysiological research allows preclinical studies to identify mechanisms of the identified pathologies that can become targets for intervention. Pyramidal cells generate broadband, persistent gamma activity37, but canonically, interneurons such as fast-spiking parvalbumin (PV) GABAergic cells, are critical to “sharpening” gamma oscillations to synchronize and shape population activity38. Loss of FMRP is associated with 1) intrinsic hyperexcitability of pyramidal neurons39 and 2) reduced activity of GABAergic circuits associated with PV cell activity40. Therefore, these are promising mechanisms of identified pathologies in FXS. Notably, increased gamma power is associated with the severity of intellectual disability in FXS, demonstrating one of the strongest correlations available of biomarkers with core features of an NDD.
Males with FXS consistently demonstrated greater gamma activity across all cortical regions than females with FXS in both vertex and network level analyses (see Figure 3c), and the association between elevated broadband gamma activity and intellectual disability was primarily driven by male participants. Residual FMRP levels in females may thus have a moderating effect on cortical hyperexcitability as measured by increases in broadband gamma. The distinctive behavioral disturbances in females might be thus more related to their abnormalities in lower frequency long distance modulation in the theta and alpha bands.
Thalamocortical modulation of neocortical hyperexcitability
Local circuit hyperexcitability is well established in FXS, but our observations of altered low-frequency modulation extends mechanistic understanding of the illness. Alpha activity, particularly in alpha2, was broadly reduced across node, region, and network comparisons in FXS. Cortical alpha rhythms are primarily generated by thalamocortical signaling with minor contributions from surrounding neocortical regions41. Thalamic alterations in FXS have been identified by structural neuroimaging: human infants with FXS display lower fractional anisotropy (FA) between thalamus and neocortex42, reduced grey matter density is found in thalamus of FXS adults27, and thalamic GABAA receptor density is reduced and correlated with intellectual disability43. However, the functional ramifications of these subcortical findings in FXS have remained poorly understood44.
Reduction in alpha power and “slowed” alpha frequency has been detected in conditions such as epilepsy, schizophrenia45, neuropathic pain, and tinnitus and is referred to as thalamocortical dysrhythmia(TCD)46. In tinnitus, an effective deafferentation of central auditory input reduces information flow to the thalamus and a shift of dominant alpha operating frequency (8-13 Hz) to the theta frequency range (5-7 Hz) is observed – a shift reminiscent of our data. These cortical regions may demonstrate elevated gamma activity as theta modulation less efficiently drives GABAA mediated lateral inhibition than alpha activity. Thus, the canonical role of alpha activity in inhibiting circuits to establish temporal windows for optimal sensory and neurocognitive processing may be diminished.
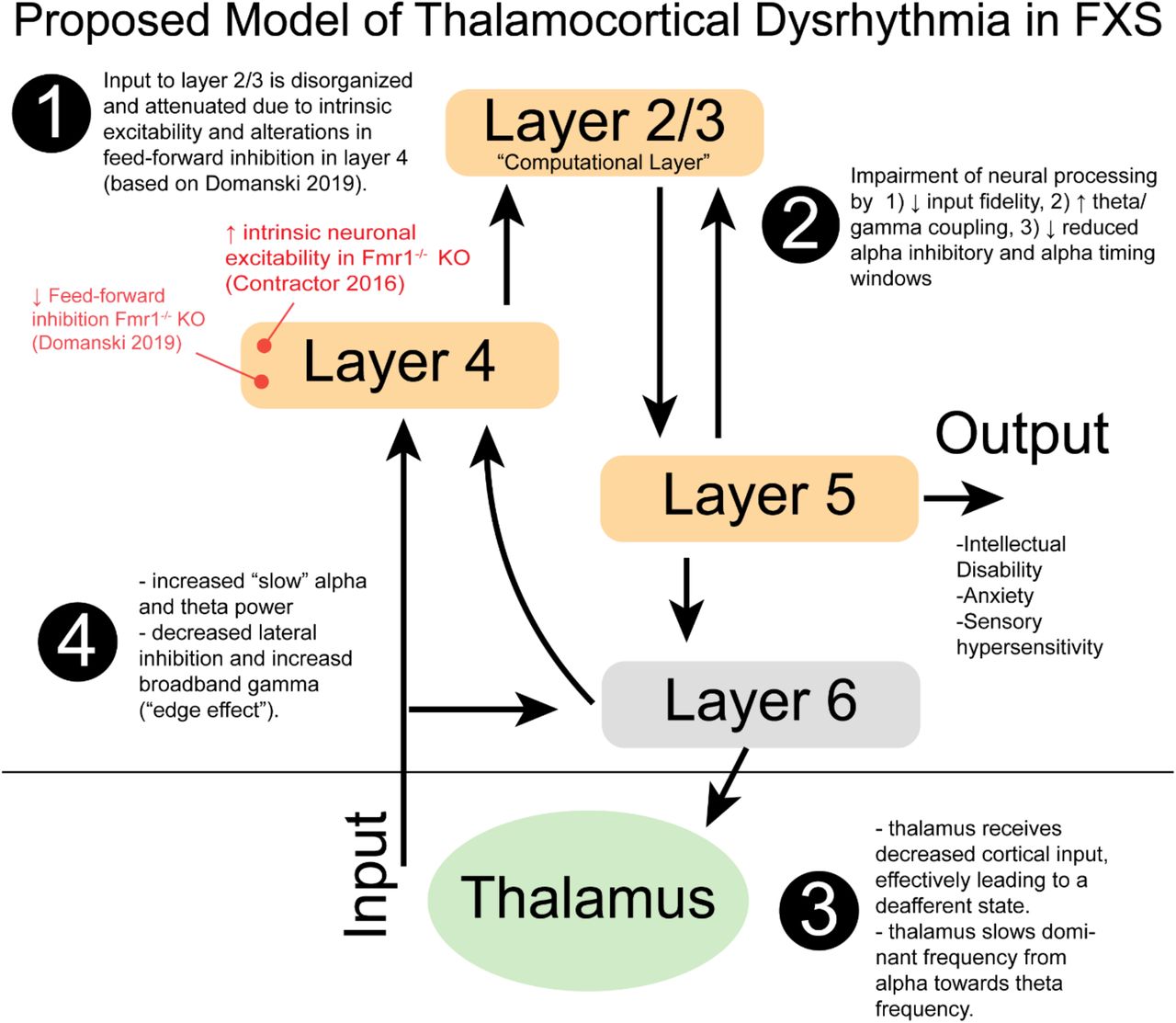
Presently, all key features of TCD were associated with FXS: 1) a leftward shift or slowing of peak alpha frequency to theta frequency, 2) an anterior redistribution of slowed alpha activity, 3) elevation of broadband gamma activity, and 4) inversion of theta and alpha power coupling with gamma activity21. Thus, in addition to well-described cell-intrinsic and local circuit hyperexcitability within the FMRP-deficient cortex, changes in thalamocortical modulation are an important additional feature contributing to increased broad band cortical excitability and intellectual disability. Fmr1-/- KO display thalamocortical mediated alterations in experience dependent plasticity of somatosensory cortex44. Importantly, the present evidence of cortical hyperexcitability at rest and prior observations of reduced gamma phase locking to auditory stimuli in FXS are conserved in the Fmr1-/- KO model11. A hypothetical model of TCD in FXS is presented in Figure 4.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hypothetical model of thalamocortical dysrhythmia (TCD) in FXS patients. In several neuropsychiatric conditions, pathological slowing of alpha oscillations is observed across M/EEG and intraoperative LFP recordings. 1) Recent study of thalamocortical circuits in Fmr1-/- KO mice observed an attenuation of complex sensory input to neocortex (layers 2/3). 2) Diminished input with other pathological changes may lead to impaired cognitive processes, 3) Thalamic feedback is effectively reduced. In the deafferent state, the dominant alpha frequency (8-13 Hz) of the thalamus is reduced to theta frequency range (5-7 Hz). 4) consequences of reduced alpha power result in additional broadband gamma, inefficient dependence on theta activity and reduced SNR of information processing. (Red, relevant Fmr1-/- KO evidence)
The interest in reducing neuronal hyperexcitability in translational models as a therapeutic strategy in FXS requires not only validated translational biomarkers but back-translational approaches to better understand mechanisms and identify novel drug targets. Importantly, these mechanisms may vary based on brain region, developmental period, circuit function, and neuronal type. In some Fmr1-/- KO circuitry, compensatory mechanisms may partially restore global homeostasis9. It is important to consider epilepsy in FXS, as despite heightened excitability, epilepsy is rare (compared to other NDDs), tends to be partial, and often resolves by adulthood or with first-line medications. Thus, while local circuit changes may disrupt specialized circuits for higher-order cognition and sensory processing47 but not to the level of representing a common cause of epilepsy. FXS subpopulations with residual FMRP activity (mosaics and female patients) may retain capacity to mount interneuron responses to “sharpen” gamma activity48 and, thus, partially spare neural functions as seen in preclinical studies49. The roles of FMRP levels in premutation cases remains to be examined.
Limitations
Although the effect of non-epileptic medications on the results cannot be ruled out, a medication naïve sample would preclude inclusion of more severely affected individuals given the high rate of medication use in FXS. Previous EEG studies of medication effects in psychiatric populations50, including our own observations in FXS14, do not suggest effects as we have observed.
Conclusion
This study of neural oscillatory activity in a relatively large sample of a monogenetic neurodevelopmental disorder provides novel evidence of hyperexcitability in functional brain networks and evidence for reduction in its thalamocortical modulation in FXS. These alterations parallel findings in preclinical models, and have clinical relevance given their association with the severity of intellectual disability. Our data from females with FXS is highly novel and provides evidence for reduced broadband gamma alterations but similar alterations in thalamocortical circuitry as in males. The alterations we observed may contribute to and maintain abnormal cortical states that reduce functional brain connectivity and regional function necessary for higher brain functions. The findings are important in establishing a robust translational strategy for developing and testing new treatments with electrophysiological biomarkers that can transfer directly from mouse model to patient studies.
Methods
Participants
The dataset included a total of 145 participants drawn from a large federally funded human neurophysiology study in FXS (National Institutes of Mental Health U54 HD082008). Exclusion criteria for FXS participants (confirmed by Southern Blot and polymerase chain reaction) included present history of unstable seizures (any treated seizure within one year) and scheduled use of benzodiazepines. Controls did not have treatment for neuropsychiatric illness as reported via clinical interview. All participants provided written informed consent (or assent as appropriate) prior to participation as approved by the institutional review board of Cincinnati Children’s Hospital Medical Center. Following blinded preprocessing, three recordings were discarded from further analysis due to excessive line-noise artifact (1 FXS, 2 controls) and one due to insufficient data due to intolerance of the EEG procedure (1 FXS). The final dataset consisted of 70 participants with a genetic diagnosis of full mutation FXS (Mean age= 20.5, SD=10; age range: 5.9-45.7; 32 females) and 71 controls (Mean age= 22.2, SD= 10.7; age range: 5.9-48.2; 30 females). Females with full mutation FXS were included in the primary analyses and effects were confirmed in supplemental analyses of male participants. Age effects were examined in each model for significant fixed effects. Thirty-five FXS patients were on antidepressants and 18 were receiving atypical antipsychotics. These and other concurrent medications were only permitted if participant was on stable dosing for at least 6 weeks.
Data Acquisition and Preprocessing
Participants were seated comfortably while watching a silent video (standardized across participants) to facilitate cooperation as in previous studies24. Dense array raw EEG data is available to the public as federally mandated at the National Database for Autism Research (NDAR). Five minutes of continuous EEG data was collected. Recordings were collected at 1000 Hz sampling rate with an EGI NetAmp 400 with a 128-channel HydroCel electrode net (Magstim/EGI, Eugene, OR). Preprocessing: All data was blinded and coded in regard to group, participant, or collection date. Data was exported in EGI raw format and imported into EEGLAB SET format in MATLAB (version 2018b, The MathWorks Inc., Natick, MA, USA). To ensure detection of neurogenic activity from the gamma band we followed best-practices to address myogenic contamination51. Raw EEG data was filtered using EEGLAB 14.1.252 with a 2 Hz high pass digital zero-phase filter and a 55 to 65-Hz notch filter (with harmonics removed up to Nyquist frequency of the original sampling rate) to remove line noise. Raw data was visually inspected by an assistant who excluded segments of data with large amount of movement artifact and interpolated bad channels (no more than 5% per subject) using spherical spline interpolation implemented in EEGLAB 14. Average number of interpolated channels was 6.2 out of 128 (SD=3.5). Data was average referenced. An artifact subspace reconstruction (ASR) approach was carried out with the “clean_rawdata” function (with default parameters) to repair data segments of artifact by applying a reconstruction mixing matrix from non-interpolated neighboring channels. The mixing matrix is computed from clean segments from within the EEG data53. Blind source separation was performed with temporal Independent Component Analysis (ICA) on each preprocessed dataset using the extended INFOMAX algorithm54, 55 with PCA rank reduction (further reduced for interpolated channels). This approach was recently validated to effectively reduce myogenic contamination from approximately 25-98 Hz18.
Resulting components were manually reviewed and categorized for eye movement/blinks, muscle movement, channel noise, or cardiac artifact based on temporospatial and spectral features and back projected to remove artifact. The average number of artifactual components removed (M=11.3; SD=1.5) was less than 10% of resultant components. Resulting non-artifactual independent components are near-independent in time course activity and resemble dipolar scalp projections and have been proposed to represent spatially coherent local field activity within a single cortical area56. Data was divided into 2-second epochs and manually reviewed for noise artifacts. Following preprocessing, an average of approximately 256 (SD=41) seconds or 128 (SD=20) epochs of artifact-free data remained for each subject.
Source Estimation and Functional Network Definition
Minimum norm estimation (MNE) is a widely adopted solution to the inverse problem in which current estimates are calculated at every spatial location in source space to minimize the total power across the cortex57. Thus, MNE models, in contrast to dipole fitting, produce uniform maps across subjects which is well-suited for group comparisons and can provide resolution comparable to magnetoencephalography (MEG)58. For each subject, the first 80-s of artifact-free time series from each of the EGI 128-channel electrodes were co-registered with an Montreal Neurological Institute (MNI) averaged ICBM152 common brain template59. The degree of accuracy and precision of EEG source localization is debated, but intracortical recordings during epileptic surgery60, surface and deep brain stimulation61, and comparisons with functional magnetic resonance imaging (fMRI)62 estimate focal localization at 1.5 cm for superficial neocortex. Thus, even with standard head models and spatial smoothing EEG is suitable for studying high-frequency brain activity in vivo clinical studies63. An open M/EEG64 was used to compute a 15,000 vertices lead-field mesh incorporating electrode distances. Noise covariance was set as an identity matrix as recommended for scalp resting EEG recordings19. Construction of L2-normed, depth-weighted MNE source model to generate a current source density (CSD) map (units: picoampere-meter) was performed in Brainstorm19 and used to reconstruct time series activations at each vertex. The Desikan-Killiany (DK) atlas was used to group vertices into 68 cortical regions, of which 44 regions were assigned within resting-state networks20. DK atlas regions were classified into five functional brain networks: default mode network (DMN), dorsal attention network (DAN), salient affective network (SAN), auditory network (AUD), and visual network (VIS) based. These dense-EEG derived networks were recently validated following examination of their dynamic properties and compared with networks identified with other neuroimaging techniques (diffusion tensor imaging, functional magnetic resonance imaging, and magnetoencephalography)2.
Spectral Power
Electrode
Estimation of power spectrum density (PSD) was computed from 108 EEG channels which contacted the scalp as a comparison for source analysis. Segmented data (2 s) from each channel was detrended, tapered with a Hanning window, and transformed into Fourier coefficients representing 0.5 Hz frequency steps. To be comparable with our previous study24, Fourier coefficients were squared to compute absolute power and divided into 7 bands of interest: delta (2–3.5Hz), theta (4–7.5 Hz), lower alpha (8–10 Hz), upper alpha (10–12.5 Hz), beta (13–30 Hz), and lower gamma (gamma1; 30–55 Hz), and higher gamma (gamma2; 65-90). We opted to split alpha band as functionally as upper alpha bands are associated with more complex cognitive processing32, 65, and lower alpha bands have been primarily associated with attentional processes including alertness, expectancy, and vigilance66. Relative power was defined as the band-specific cumulative absolute power divided by the total power across all defined bands, and then averaged over available trials. Average dominant frequency (i.e. alpha peak) was determined by the “findpeak” function in MATLAB to identify frequency of the maximum absolute logarithmic power between 6-14 Hz from each channel’s spectrogram67.
Source Analysis
Welch’s method was used to estimate spectral power from CSD between 2 to 90 Hz at each vertex with a sliding Hamming window overlapping at 50%. Relative power was calculated by normalizing spectral power per subject and dividing total power from 2-90 Hz and averaged across identical frequency bands as scalp-EEG analysis. For each subject, PSD results were projected onto the 15,000 vertices cortical source model. To facilitate group comparisons, we used a circularly symmetric Gaussian smoothing kernel with a full width half maximum (FWHM) size of 3 mm68 across all vertices.
Clinical Measures
Stanford-Binet Intelligence Scale 5th Ed. (SBS)69 was conducted by trained clinicians in both FXS and control participants. Due to floor effects, deviation IQ scores70 were computed to capture variability in cognitive functioning. Assessments were completed by the primary caregivers for FXS patients including the Social Communication Questionnaire (SCQ)71, Anxiety, Depression, and Mood Scale (ADAMS)72, Woodcock-Johnson III Tests of Cognitive Abilities, Auditory Attention subscale (WJ3)73.
Statistics
Software: Statistical analysis was performed with MATLAB 2018b (MathWorks, Natick, MA, USA), SAS 9.4 (SAS Institute Inc., Cary, NC, USA), and R (4.0, Vienna, Austria). Power and Sample Size: Differences in gamma1 power in FXS compared to controls in previous studies have effect sizes from .63 to 1.75, similar to effect sizes in prior studies of N1 amplitudes in FXS6, 14, 24, 74. Based on these effect sizes, comparing 70 FXS patients (50% males) and 70 TD controls provides power to detect the primary EEG outcome with approximately power > .90 (using an omnibus F-test with an alpha of .05). In line with reproducible research guidelines, scripts for generation of figures and tables are available upon request.
Group comparisons
Electrode-based spectral power comparison: Cluster-based permutation analysis75 was used to identify significant differences between FXS and controls in relative power electrode by electrode between frequency bands. Overall alpha was set at .05 / 7 (adj. p.<.007) to account for multiple band comparisons (effective alpha for each tail .025). Source-localized spectral power comparison: Group-level statistical (t-statistic) maps were assessed by Monte-Carlo permutation (2000) independent two-tailed t-tests (alpha set at .025 per tail) using the ‘ft_sourcestatistics’ function in FieldTrip76 and threshold at P<.05. The resulting p values were globally corrected by a false discovery rate (FDR) of 5% applied over the signals and frequency band dimensions77.
Node and Network Comparisons
Log-transformed relative power differences were evaluated with generalized linear mixed models via the GLIMMIX procedure78 in SAS 9.4 (SAS Institute Inc., Cary, NC, USA) in which random effect was subject and independent variable varied based on model. Vertex-by-vertex model: LME included fixed effects of group (FXS vs. control), node (68 atlas nodes), and seven frequency bands on log-transformed relative power. RSNs: Group: Model included fixed effects of Group (FXS vs. control), RSN (DMN, VIS, DAN, SAN, vs AUD), seven Frequency bands, and DK atlas nodes. Sex Differences: The final fitted model included subject as the random effect and fixed effects of Group (FXS vs. control), Sex (M vs. F), RSN (DMN, VIS, DAN, SAN, vs AUD), and seven Frequency bands. In this model atlas nodes within each RSN were treated as replicates. Different link functions and intra-subject covariance structures were examined. Plots based on the studentized residuals were examined in each case.
Alpha-shift Distribution
An LME was conducted to assess CFC differences in which the random effect was subject and with fixed effects of group (FXS vs. control), sex (male vs. female), and lower CFC band (theta vs. alpha) on gamma1 amplitude-amplitude coupling (AAC). We examined within group alpha-shift changes along the occipital-frontal axis using an LME to model log-transformed αshift with fixed effects of group (FXS, Control), sex (M vs. F), and occipital-frontal axis region (occipital, parietal, central, and frontal; composed of atlas nodes) with the final fitted model including subject as the random effect and controlling for age.
Cross frequency amplitude coupling
To examine potential dependence between low-frequency activity and high-frequency activity, we evaluated cross-frequency amplitude-amplitude coupling (AAC) consistent with previous reporting24 with low-frequency band candidates, theta and alpha band (8-12.5 Hz), with gamma1 or gamma2. CSD time series from each DMN node were segmented into 2-s epochs. Relative powers per epoch from the selected band candidates contributed to the Spearman Correlation for all DMN nodes. Normalization via Fisher’s Z-transform was applied for groupwise comparisons.
Correlation analysis
As a successive step, frequency bands of significance were linearly correlated with clinical and behavioral measures. Primary Analysis: Shapiro-Wilk’s normality test was performed on variables to assess suitability for either Spearman’s rank-order or Pearson’s correlation test. A priori hypotheses for high-frequency bands (beta, gamma1, and gamma2) and low-frequency bands (theta, alpha1, and alpha2) with clinical variables were assessed with correlation tests with p values adjusted by FDR for multiple test iterations and partial correlations were used to adjust all correlations for age.
Data Availability
EEG data is available to the public as federally mandated at the National Database for Autism Research (NDAR). Analysis code is in preparation for upload into public repositories and is available on request.
Acknowledgments
We thank the participants and families who participated in this study. We would also like to thank Nicole Friedman, Michael Hong, Danielle Chin, and Janna Guilfoyle who assisted with project. The present study was federally funded by National Institutes of Health (NIH) Fragile X Center (U54HD082008).
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵



