
Abstract
Background Mass vaccination against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is on-going in the United States with vaccines approved for emergency use by the FDA. Each vaccine can induce spike-specific antibodies (Abs) with virus-neutralizing activities; however, the Fc-mediated Ab activities have received little attention. Moreover, while plasma/serum Abs are commonly studied, scant information is available about Abs in the respiratory mucosa, the site of SARS-CoV-2 transmission.
Methods Plasma and saliva were collected from mRNA vaccine recipients and convalescent coronavirus disease 2019 (COVID-19) patients. Antigen-specific total Ig and Ig isotypes were measured. In addition to virus neutralization, Fc-mediated activities were investigated, including antibody-dependent cellular phagocytosis (ADCP) and complement deposition.
Results Similar to infection, vaccination stimulated spike-specific Ab responses detected in plasma and saliva, with IgG1 as the dominant isotype. Interestingly, vaccination produced greater IgG2, IgG3, and IgG4 responses and higher ratios of (IgG1+IgG3)/(IgG2+IgG4) than infection. Moreover, while plasma neutralization and ADCP potencies were comparable in vaccinated and convalescent individuals, vaccine-induced plasma Abs elicited stronger complement binding and activation.
Conclusion Compared with natural infection, mRNA vaccines induced a greater array of IgG subtypes against spike in saliva and plasma. The vaccine-induced Abs were also more potent in mediating complement activation.
Background
In the USA, three coronavirus disease-2019 (COVID-19) vaccines have received an authorization for emergency use from the FDA: two messenger RNA (mRNA) vaccines from Pfizer-BioNTech (BNT162b2) and Moderna (mRNA-1273), and one adenovirus-vectored vaccine from Johnson & Johnson/Janssen (Ad26.CoV2.S). The two mRNA-based vaccines reported >90% efficacy in preventing symptomatic SARS-CoV-2 infection [1,2], while the efficacy of Ad26.CoV2.S was ∼70% [3]. All three vaccines were designed to induce antibodies (Abs) targeting SARS-CoV-2 spike [4,5], a membrane-anchored protein on the viral surface which contains the receptor-binding domain (RBD) necessary for binding and entry into the host cells [6–8].
While the primary antiviral function of Abs is to neutralize the virions, Abs also mediate non-neutralizing effector functions via their Fc fragments. In the case of SARS-CoV-2, the binding of anti-spike Abs to virions, infected cells, or soluble spike proteins creates immune complexes capable of engaging Fc receptors (FcRs) or complement via the Abs’ Fc fragments [9,10]. The FcR engagement triggers a cascade of intracellular signals critical for Fc-mediated activities, such as Ab-dependent cellular phagocytosis (ADCP) and Ab-dependent cellular cytotoxicity (ADCC). The binding of C1q, the first component in the classical complement pathway, to Fc fragments on immune complexes activates the downstream complement cascade, resulting in the deposition of C3 and C4 degradation products and leading to the generation of C5 convertase and the assembly of the membrane-attack complex responsible for initiating complement-mediated lysis. Complement-opsonized immune complexes also interact with complement receptors on leukocytes to trigger effector functions, such as complement-dependent cell-mediated phagocytosis and cytotoxicity [11,12].
The Ab affinity for FcR and complement is determined by their immunoglobulin (Ig) isotypes. Each of the Ig isotypes interacts with distinct FcRs, categorized into type I and type II [9,13]. The canonical type I FcRs for IgG include FcγRI (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16A), and FcγRIIIB (CD16B), which differ in cell distribution, affinity for IgG subtypes, and functional effects. Among IgG subtypes, IgG1 and IgG3 have the strongest affinity for the activating FcγRI, FcγRIIA, and FcγRIIIA [14]. On macrophages, the engagement of FcγRI and FcγRIIA stimulates ADCP. On NK cells and some macrophages, FcγRIIIA serves as the key receptor for inducing ADCC [15,16]. Each Ab isotype also activates the complement system with varying potency in the following order: IgM=IgG3>IgG1>IgG2=IgA, while IgG4, IgD, and IgE do not engage complement [17].
The COVID-19 vaccines used for mass immunization demonstrate the capacity to elicit neutralizing Ab responses, but the full properties of the elicited Abs are not yet known [2,18–21]. In particular, Fc-mediated activities and the associated Ig isotypes elicited after vaccination have not been studied. Moreover, while SARS-CoV-2-specific Ab profiles from convalescent individuals have been shown to be similar in plasma and saliva [22], the Ab profiles in the respiratory mucosa, the initial site of SARS-CoV-2 infection, from vaccinated individuals remains to be investigated. In this study, using a multiplex bead assay developed in our previous study [23], we analyzed the spike- and RBD-specific Ig isotypes in paired plasma/saliva specimens from seven mRNA vaccine recipients in comparison with eight COVID-19-positive convalescent donors and four COVID-19-negative individuals. Additionally, the vaccine-induced plasma Abs were examined for Fab-mediated virus neutralization and Fc-mediated activities (ADCP, C1q binding and C3d deposition). This study provides the first evidence that SARS-CoV-2 immunization induces a greater breadth of spike-specific Ig isotypes and more potent complement binding capacities than natural infection.
Methods
Human specimens
Plasma and saliva specimens were obtained from volunteers enrolled in IRB-approved protocols at the Icahn School of Medicine at Mount Sinai (IRB#17-00060, IRB#19-01243) and the James J. Peter Veterans Affairs Medical Center (IRB#BAN-1604): RN#1, RN#4 and RV#1-5 after immunization; RP#2-5, 7, 12-14 after infection; contemporaneous non-vaccinated COVID-19-negative and pre-pandemic plasma samples. The clinical data are summarized in the Supplementary Tables 1 (vaccinated individuals) and 2 (convalescent individuals). All participants provided written consent. All samples were heat-inactivated before use.
Multiplex bead Ab binding assay
Measurement of total Ig and Ig isotypes to SARS-CoV-2 antigen-coupled beads was performed as described [23]. Complement deposition onto plasma and saliva Abs reactive with spike and RBD were measured according to [26] with modifications. For the C1q assay, beads with spike-Ab or RBD-Ab complexes were incubated with C1q Component from Human Serum (Sigma, #C1740) for 1 hour at room temperature, followed by an anti-C1q-PE antibody (Santa Cruz, #sc-53544 PE). For the C3d assay, Complement Sera Human (33.3%, Sigma, #S1764) was added to the beads for 1 hour at 37°C, followed by a biotinylated monoclonal anti-C3d antibody (Quidel, #A702).
Virus neutralization
VSV-based SARS-CoV-2 pseudoviruses were used in neutralization assays as described [23,27].
ADCP
ADCP assays were performed using a reported protocol [28] with some modifications. FluoSpheres carboxylate-modified microspheres (Thermo Fisher, #F8823) were coupled with SARS-CoV-2 spike protein using the xMAP Antibody Coupling Kit (5 µg protein/∼36.4⨯109 beads, Luminex #40-50016). Spike-conjugated microspheres were incubated with diluted plasma for 2 hours at 37°C in the dark. After washing and centrifugation (2,000 g, 10 minutes), the beads (∼3⨯108 beads, 10 µL/well) were incubated with THP-1 cells (0.25×105 cells, 200 µL/well) for 16 hours. The samples were analyzed on an Attune NxT flow cytometer (Thermo Fisher, #A24858). Data analysis was performed using FCS Express 7 Research Edition (De Novo Software).
Statistical analysis
Statistical analyses were performed as designated in the figure legends using GraphPad Prism 8 (GraphPad Software, San Diego, CA).
Results
COVID-19 mRNA vaccines induce Ab responses to SARS-CoV-2 spike, RBD, S1 and S2 antigens
Paired plasma and saliva specimens from seven healthy recipients of COVID-19 mRNA vaccines were examined for total Ig and different Ig isotypes against the SARS-CoV-2 antigens. Two individuals received the Pfizer vaccine, five received the Moderna vaccine, and samples were collected 15-37 days after the second vaccine dose (Supplemental Table 1). Four COVID-19-negative non-vaccinated contemporaneous donors were tested for comparison and used to establish the cut-off values.
The plasma specimens were titrated for total Ig against spike, RBD, S1, S2, and nucleoprotein antigens, with bovine serum albumin (BSA) as a control antigen (Fig. 1a). All vaccinated subjects exhibited titrating amounts of plasma Ig against spike, RBD, S1 and S2, above the cut-off levels. As expected, no reactivity was observed against the nucleoprotein, although one individual had weak reactivity at the lowest dilution, reaching slightly above the cut-off.
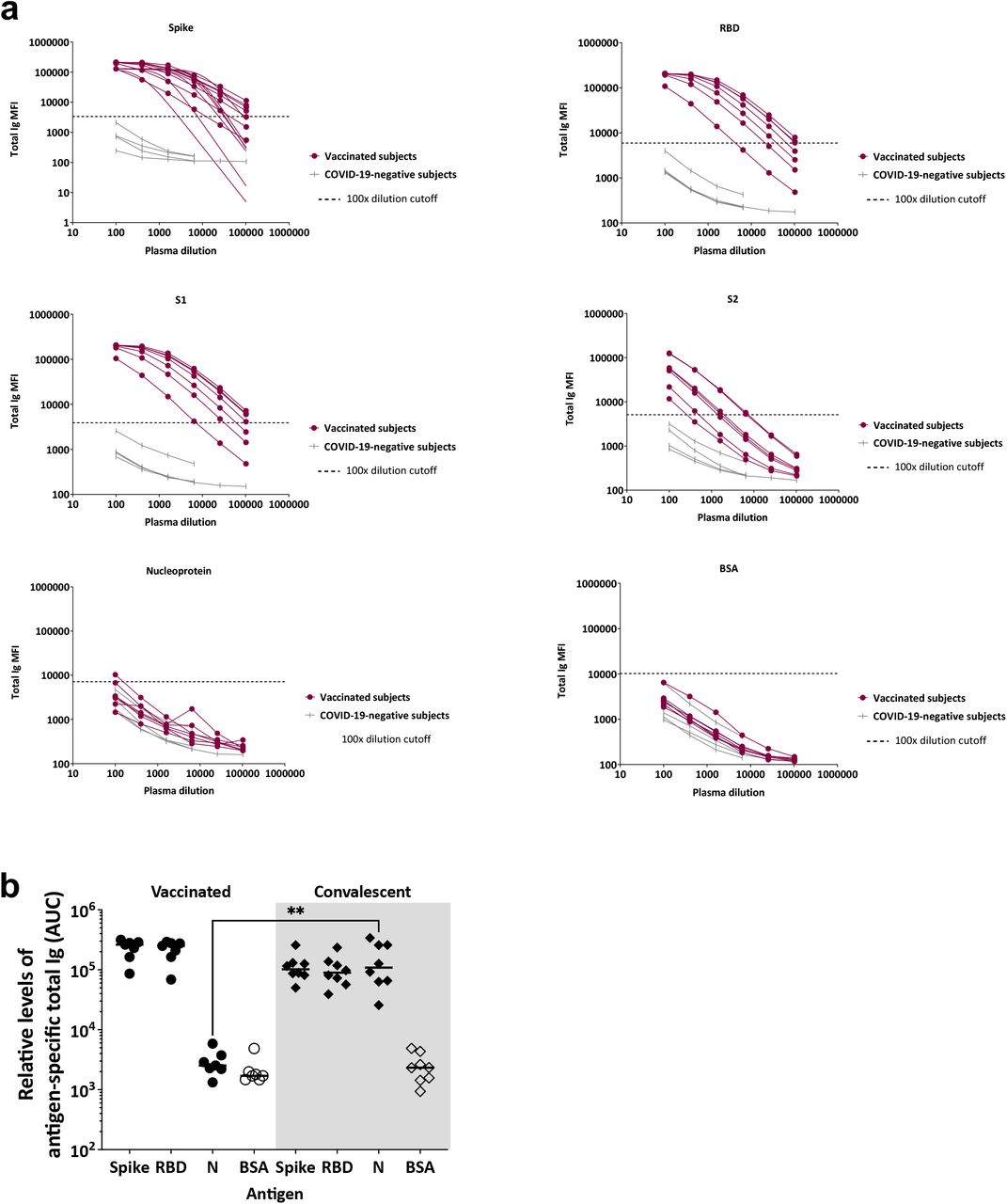
(a) Titration curves are shown for total Ig against spike, RBD, S1, S2, nucleoprotein, and BSA in plasma specimens from seven vaccinated and four COVID-19-negative donors. Specimens were diluted at 4-fold dilutions from 1:100 to 1:6,400 or 102,400. The dotted gray lines indicated the cut-off calculated as the mean + 3SD of a 1:100 dilution of the four COVID-19 negative specimens. All data were generated using the multiplex bead antibody binding assay and plotted as mean fluorescent intensity (MFI). (b) Relative levels of plasma total Ig against spike, RBD, nucleoprotein (N), and BSA, as measured by area under the curves (AUC), in specimens from seven vaccinated donors and eight convalescent COVID-19 individuals tested in parallel. Statistical significance was calculated using a Kruskal-Wallis test.
Comparison with plasma samples from eight convalescent COVID-19-positive specimens (collected 40-256 days post-symptom onset, Supplemental Table 2) showed differential Ig responses against nucleoprotein. The area under the curve (AUCs) values of vaccinated and convalescent specimens were not significantly different, although the median AUCs of the vaccinated group were 2.6-fold (for spike) and 2.8-fold (for RBD) higher than those of the convalescent group (Fig. 1b).
COVID-19 mRNA vaccines induce varying levels of Ig isotypes against SARS-CoV-2 spike
The plasma specimens were then tested for antigen-specific total Ig, IgM, IgG1-4, IgA1 and IgA2 at a 1:200 dilution (Fig. 2a). The percentage of responders for each isotype was determined using the cut-off values calculated as mean+3 standard deviation (SD) of 12 pre-pandemic samples reported previously [23]. All vaccinated plasma specimens had high levels of anti-spike and anti-RBD total Ig and IgG1. A significant proportion (40-86%) of specimens also displayed IgG2-4 Abs against spike and RBD, although the levels were low. Notably, 100% and 71% of individuals produced IgA1 Abs against spike and RBD, respectively, while 14% and 57% exhibited spike- and RBD-specific IgA2. In addition, 71% generated spike-specific IgM at varying levels. Only 14% had RBD-specific IgM mainly due to higher background, consistent with our previous findings [23]. None of the four control specimens showed Ig isotype reactivity with spike or RBD.

(a) Total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1 and IgA2 levels against spike and RBD were measured in plasma specimens from seven vaccinated and four COVID-19-negative individuals. The black line represents the median for each group. The percentages of responders above the cut-off for each spike- or RBD-specific Ig isotype are indicated. (b) Simple linear regression of spike-specific or RBD-specific plasma total Ig levels versus IgM, IgG1, IgG2, IgG3, IgG4, IgA1, and IgA2 levels from the seven vaccinated individuals. (c) Spike- or RBD-specific ratios of IgG1+IgG3 MFIs over IgG2+IgG4 MFIs for the vaccinated and convalescent groups. Statistical significance was calculated using a Mann-Whitney test.
Linear regression analysis was performed to assess the relative contribution of each isotype to the total Ig responses to spike and RBD (Fig. 2b). IgG1 had the highest and most significant r2 values, indicating that IgG1 is the major isotype induced by COVID-19 mRNA vaccines. In addition, IgG3 Abs demonstrated a significant contribution, with an r2 of 0.70 for spike and 0.64 for RBD with p values <0.05 for both.
Comparison of spike- and RBD-specific Ig isotypes generated by vaccination versus infection
Linear regression data of plasma specimens from the seven vaccinated and eight convalescent subjects tested in this study (Supplemental Fig. 1) were comparable to those from a previously reported cohort of 29 convalescent subjects [23], demonstrating that IgG1 was the dominant isotype induced after vaccination or infection. However, the association with IgG3 Abs was stronger in plasma from vaccinated versus convalescent donors, where a significant association was observed with RBD-specific IgG3 only (Supplemental Fig. 1, [23]).
Further comparison of Ig isotypes elicited by vaccination versus infection demonstrated that the percentage of IgM responders in the vaccinated group was lower than those observed in our past study of 29 convalescent individuals [23] and the eight convalescent donors examined in parallel in this study (88% for spike and 75% for RBD, Supplemental Fig. 1). In contrast, the percentages of IgG2, IgG3, and IgG4 responders were higher (Supplemental Fig. 1), indicating that vaccination induces a broader range of IgG subtypes. A heatmap is shown in Fig. 3a to illustrate these differences.

Heatmaps to show the relative levels of antibody binding, neutralization and Fc-mediated activities in (a) plasma and (b) saliva specimens from seven vaccinated donors, eight COVID-19 convalescent patients, and four COVID-19-negative controls. Y-axis represents each individual and X-axis represents each Ab binding or activity studied. Data were normalized by dividing each individual data by the highest value of the group.
Interestingly, the ratios of IgG1+IgG3 mean fluorescence intensity (MFIs) to IgG2+IgG4 MFIs were higher in the plasma of the seven vaccinated individuals than in those of the eight convalescent individuals (p<0.05) (Fig. 2c), further distinguishing the two groups. A similar pattern was seen when comparing these seven vaccinated specimens with the previously studied cohort of 29 convalescent plasma specimens from <8 weeks post-symptom onset: the median ratios of spike-specific (IgG1+IgG3)/(IgG2+IgG4) were 133 and 46 for the vaccinated and convalescent groups, respectively (p=0.0019). For RBD-specific Abs, the median ratio of the vaccinated group was also two times higher (p=0.06). These data demonstrate that, while vaccine-induced Ab responses are comprised mainly of spike- and RBD-specific IgM, IgG1, and IgA1, vaccination elicited higher proportions of IgG1 and IgG3 over IgG2 and IgG4 than natural infection.
Vaccine-induced IgG1 and IgG3 against spike and RBD contribute the most to virus neutralization activities
To test the neutralizing activity of vaccine-induced plasma Abs, specimens from seven vaccinated individuals and two non-vaccinated COVID-19-negative individuals were examined in a neutralization assay using a VSVΔG pseudovirus bearing the SARS-CoV-2 spike protein. All vaccinated samples attained 90% neutralization, while none of the COVID-19-negative samples did (Fig. 4a). The IC50 titers attained after vaccination were similar to those of convalescent samples collected <8 weeks after symptom onset [23] or at later time points (>186 days) (Fig. 4b, Fig. 3a).
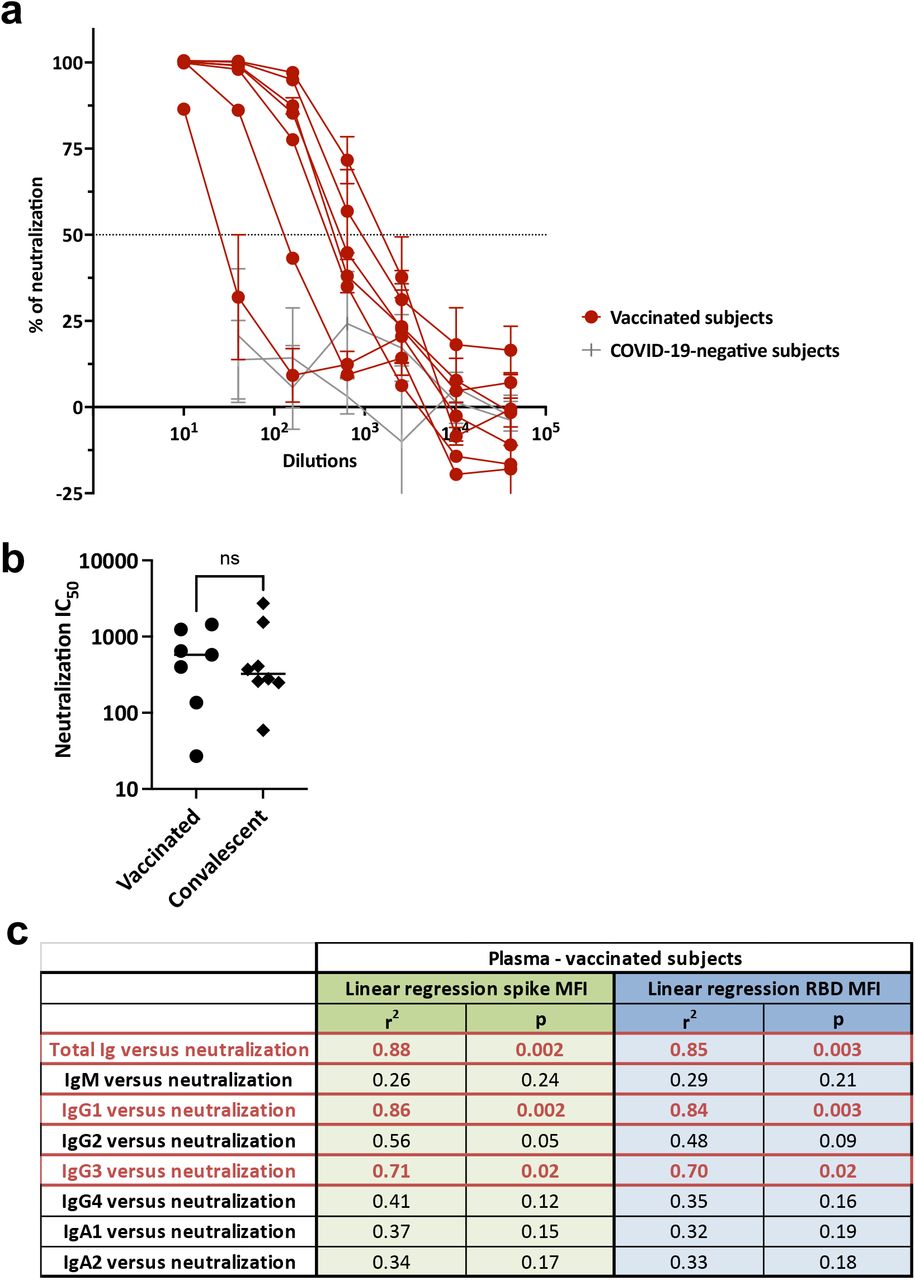
(a) Neutralization of SARS-CoV2 pseudovirus by plasma specimens from seven vaccinated individuals and two COVID-19-negative individuals. The dotted line indicates 50% neutralization. (b) Neutralization IC50 titers of plasma from seven vaccinated compared to eight convalescent donors. Statistical significance was calculated using a Mann-Whitney test. (c) Simple linear regression of neutralization IC50 titers versus spike- or RBD-specific total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1, and IgA2 levels from the seven vaccinated individuals.
Linear regression analyses were performed to analyze the contribution of each isotype to the neutralization activities (Fig. 4c). Similar to our past data with convalescent plasma [23], high r2 values (0.84-0.85) and significant p values were observed with total Ig and IgG1. Spike- and RBD-specific IgG3 Abs also demonstrated a significant association with neutralization, with r2 values of 0.71 and 0.70, respectively. Interestingly, IgM did not show any association. This result is contrary to our observation in convalescent individuals [23] but is in line with lower IgM responses detected in these vaccinees after the second vaccine dose. The association between IgM and neutralization was also not found in convalescent samples collected at late time points (Supplemental Fig. 2), as the IgM response was lower in these samples compared to specimens collected <8 weeks after symptom onset [23].
COVID-19 mRNA vaccines induce ADCP activities
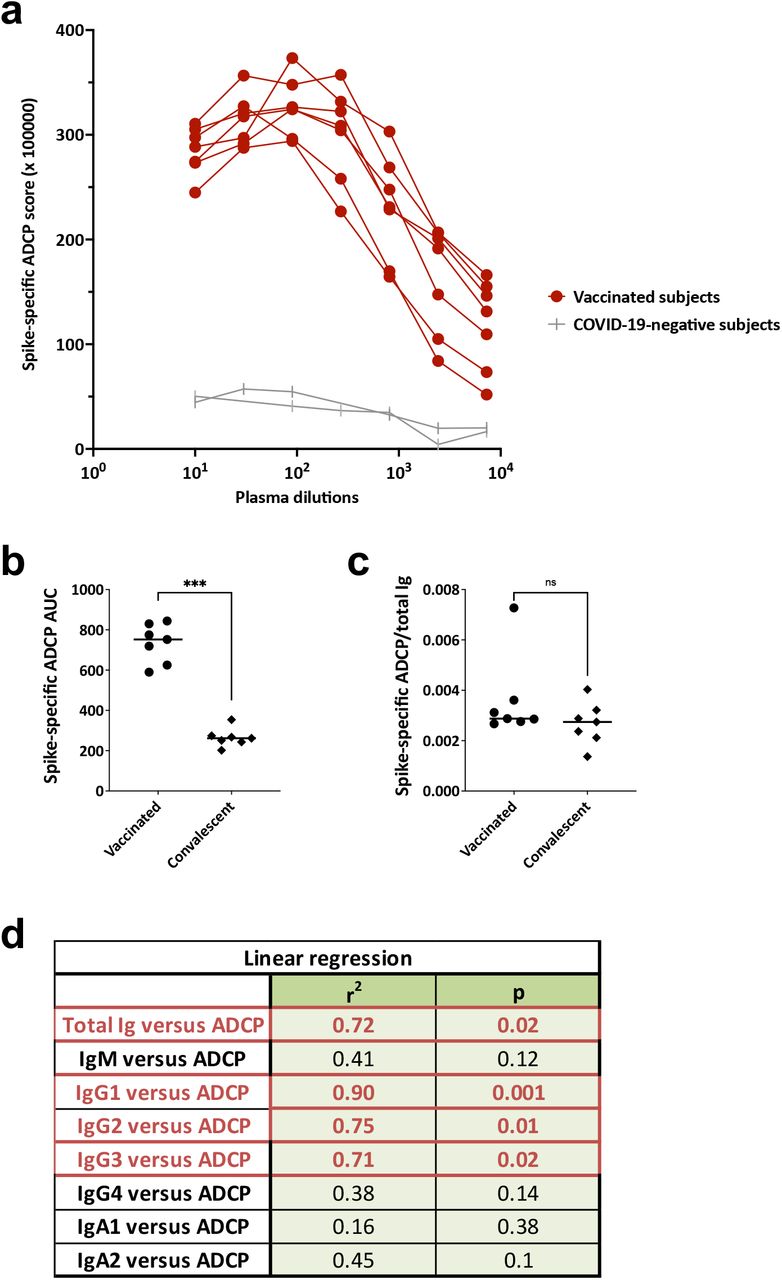
Because Ig isotypes are key determinants of Fc functions, we examined the Fc-mediated Ab activities in plasma specimens from the seven vaccinated donors in comparison with convalescent plasma and negative controls. We evaluated 1) spike-specific ADCP using THP-1 phagocytes and spike-coated fluorescent beads and 2) complement activation based on C1q and C3d deposition on spike-Ab and RBD-Ab complexes.
All specimens from vaccinated subjects displayed ADCP activities above the negative control (Fig. 5a), similar to those seen in convalescent specimens (Supplemental Fig. 3, Fig. 3a). The ADCP levels were higher for samples of vaccinated donors versus convalescent donors (Fig. 5b, Fig. 3a), corresponding to an ∼2.6-fold difference in spike-binding total Ig median levels (Fig. 1a). To compare the ADCP capacity of plasma Abs from vaccinated versus convalescent donors independent of the spike-binding Ig levels, we calculated the AUC ratios of spike-specific ADCP over the spike-binding total Ig. The ADCP capacity of Abs induced by vaccination versus infection was equivalent (Fig. 5c), demonstrating that mRNA vaccines and SARS-CoV-2 infection induce plasma Abs with comparable ADCP potency.

(a) Spike-specific ADCP activities in plasma specimens from seven vaccinated individuals and two COVID-19-negative individuals. (b) ADCP AUC for the seven vaccinated and seven COVID-19 convalescent donors. Statistical significance was calculated using a Mann-Whitney test. (c) ADCP potency was calculated as ADCP AUC/total Ig AUC for the seven vaccinated and seven COVID-19 convalescent donors. Statistical significance was calculated using a Mann-Whitney test. (d) Simple linear regression of ADCP AUC versus spike-specific total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1, and IgA2 levels in the seven vaccinated individuals.
To assess the contribution of each isotype to ADCP in the plasma of vaccinated donors, regression analyses showed the highest r2 value and the most significant p value for spike-specific IgG1 Abs (Fig. 5d). Interestingly, spike-specific IgG2 and IgG3 Abs also demonstrated significant associations with ADCP, with r2 and p values comparable to those of total Ig but lower than those of IgG1. The ADCP association with IgG1 and IgG2 Abs, but not IgG3 Abs, was also observed with convalescent plasma (Supplemental Fig. 3).
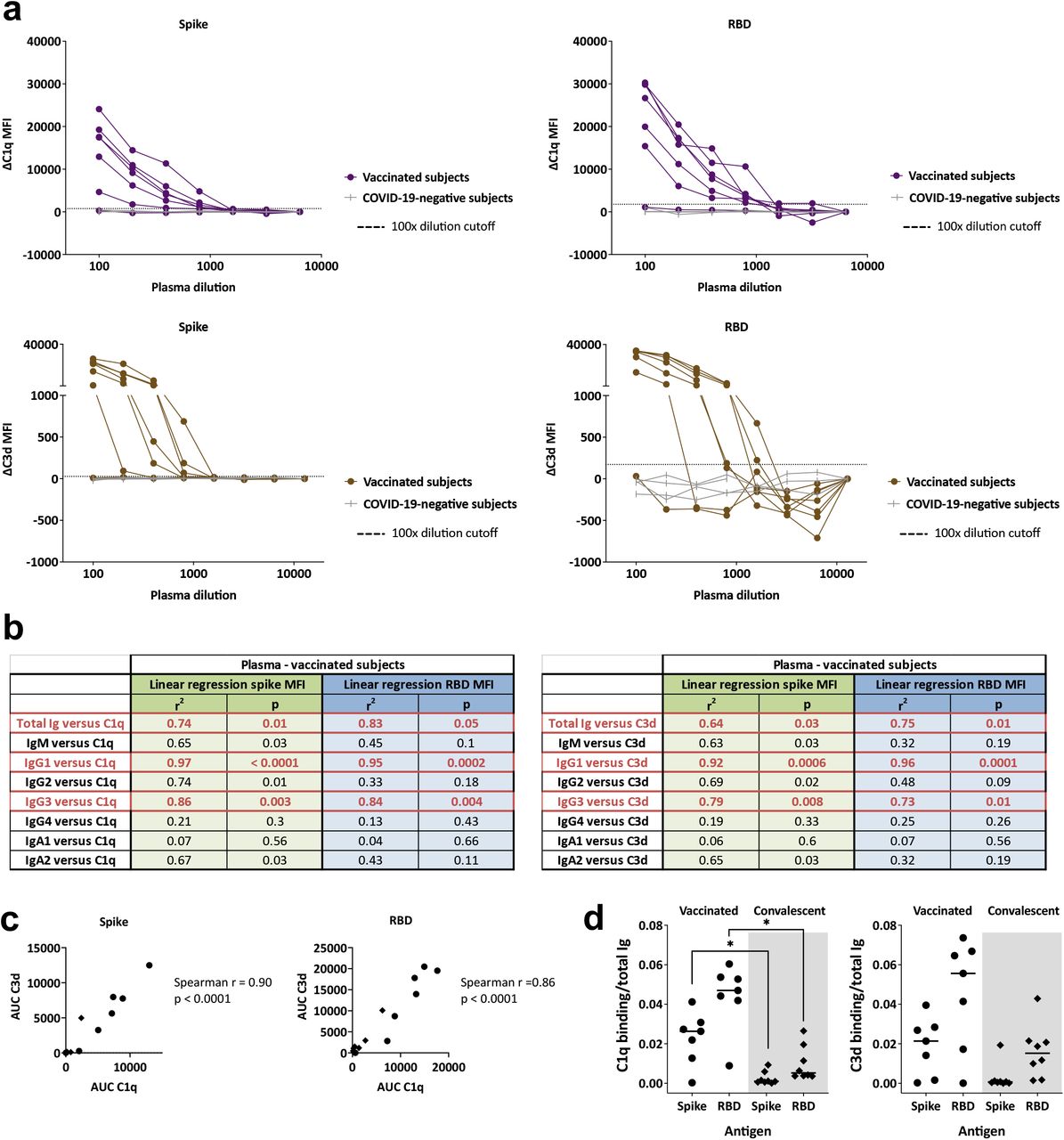
COVID-19 mRNA vaccines induce Abs with greater C1q and C3d deposition activities
When we examined C1q and C3d deposition, a high amount of variability was observed among the seven vaccinated donors. One plasma sample did not show any C1q or C3d binding to spike- or RBD-binding Abs (Fig. 6a), and the remaining samples exhibited a range of C1q and C3d binding levels. Regression analyses demonstrated that, among the spike- and RBD-binding Ig isotypes tested, IgG1 and IgG3 Abs contributed the most to the complement binding activities of the vaccinated plasma specimens (Fig. 6b). This is consistent with the greater potency of IgG1 and IgG3 Abs, as compared with IgG2 and IgG4 Abs, to activate the classical complement cascade [23]. C1q and C3d deposition was also detected with convalescent plasma samples, albeit at lower levels (Fig. 3a, Supplemental Fig. 4). The binding levels of C1q and C3d correlated with each other for spike-and RBD-specific Abs in vaccinated and convalescent donors (Fig. 6c).
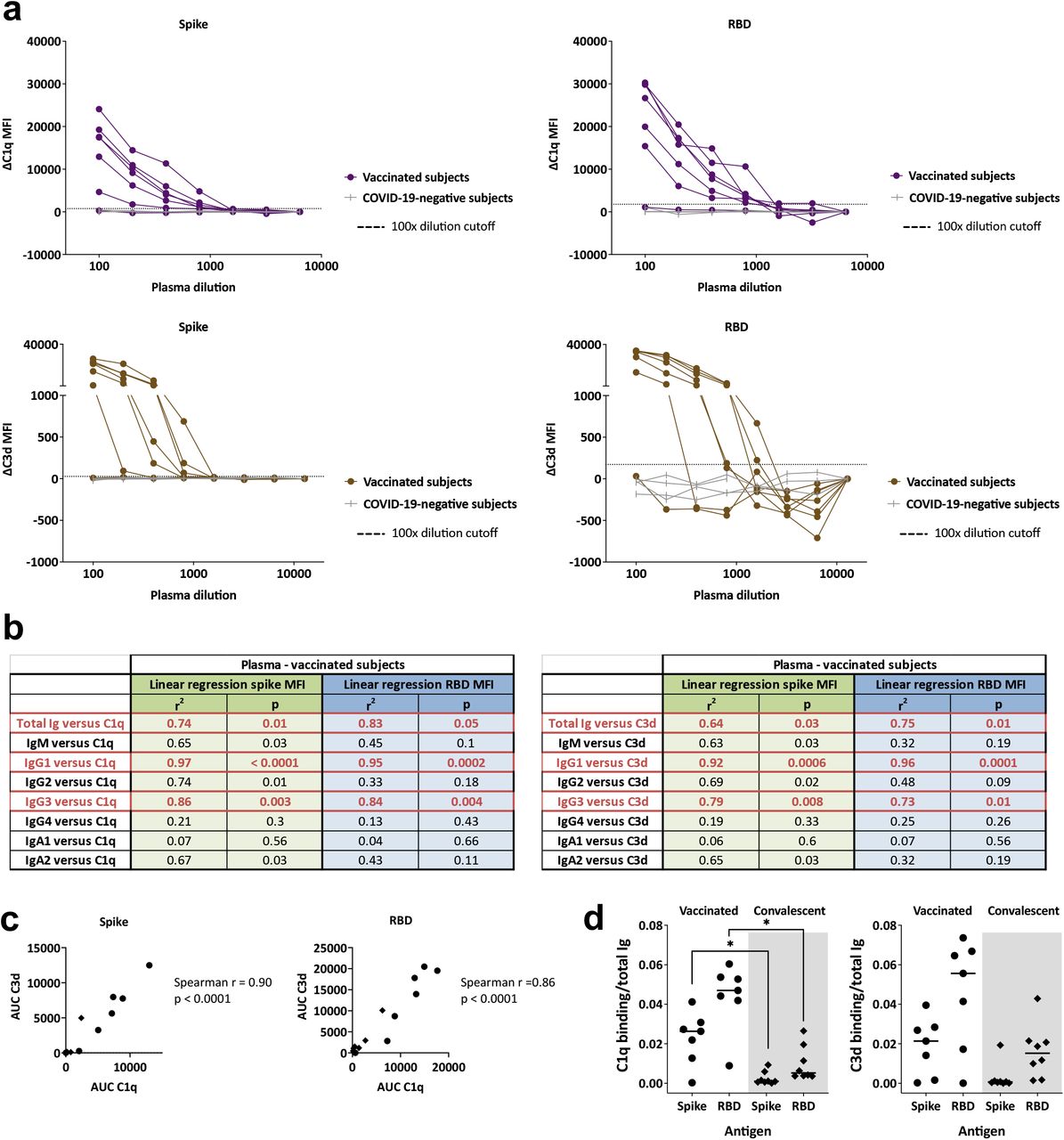
(a) C1q or C3d binding to spike- or RBD-specific Abs in plasma specimens from seven vaccinated individuals and four COVID-19-negative controls. Specimens were diluted at 2-fold dilutions from 1:100 to 1:6,400 or 12,800. The dotted line represents the 100x dilution cut-off calculated as mean of the four control specimens + 3SD. ΔC1q and ΔC3d MFI values were calculated by subtracting background MFI from each assay. (b) Simple linear regression of C1q or C3d AUC values versus spike-specific total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1, and IgA2 levels from the seven vaccinated individuals. (c) Spearman correlation between spike- or RBD-specific Ab C1q AUC and C3d AUC for plasma specimens from the seven vaccinated (circles) and eight convalescent (diamonds) donors. (d) Complement binding and activation potencies of vaccine-versus infection-induced Abs were calculated by dividing the C1q or C3d AUC with spike-specific Ig AUC for each of the seven vaccinated and eight convalescent individuals. Statistical significance was calculated using a Kruskal-Wallis test.
To compare the complement binding capacity of vaccine-versus infection-induced Abs, the ratios of C1q or C3d binding AUCs over total Ig AUCs were calculated. C1q and C3d binding capacity was higher for vaccine-induced Abs, although only the difference in C1q binding reached statistical significance (Fig. 6d). Hence, the spike- and RBD-specific Abs induced by vaccination exhibited a greater complement activation potency than the infection-induced counterparts.
We summarize the isotypes and functions of plasma Abs induced after vaccination versus natural infection in Fig. 3a. The heatmap clearly shows a greater breadth of Ig isotypes against spike and RBD in the plasma of vaccinated versus convalescent subjects. This was accompanied by more potent Fc-dependent activities especially complement activation, even though neutralization potency was indistinguishable.
COVID-19 mRNA vaccines induce spike-specific Ab responses in saliva
Since SARS-CoV-2 is a respiratory virus, we asked whether the COVID-19 mRNA vaccines also elicited Ab responses against the spike antigen in the upper respiratory mucosa where the Abs would serve as the frontline defense against this virus. To this end, we examined the levels of antigen-specific total Ig and Ig isotypes in saliva, as a proxy for oral and upper respiratory mucosal secretion, following vaccination versus natural infection. Saliva specimens were obtained from the same seven vaccinated and eight convalescent donors at the time of blood collection and compared to four non-vaccinated COVID-19 negative saliva specimens.
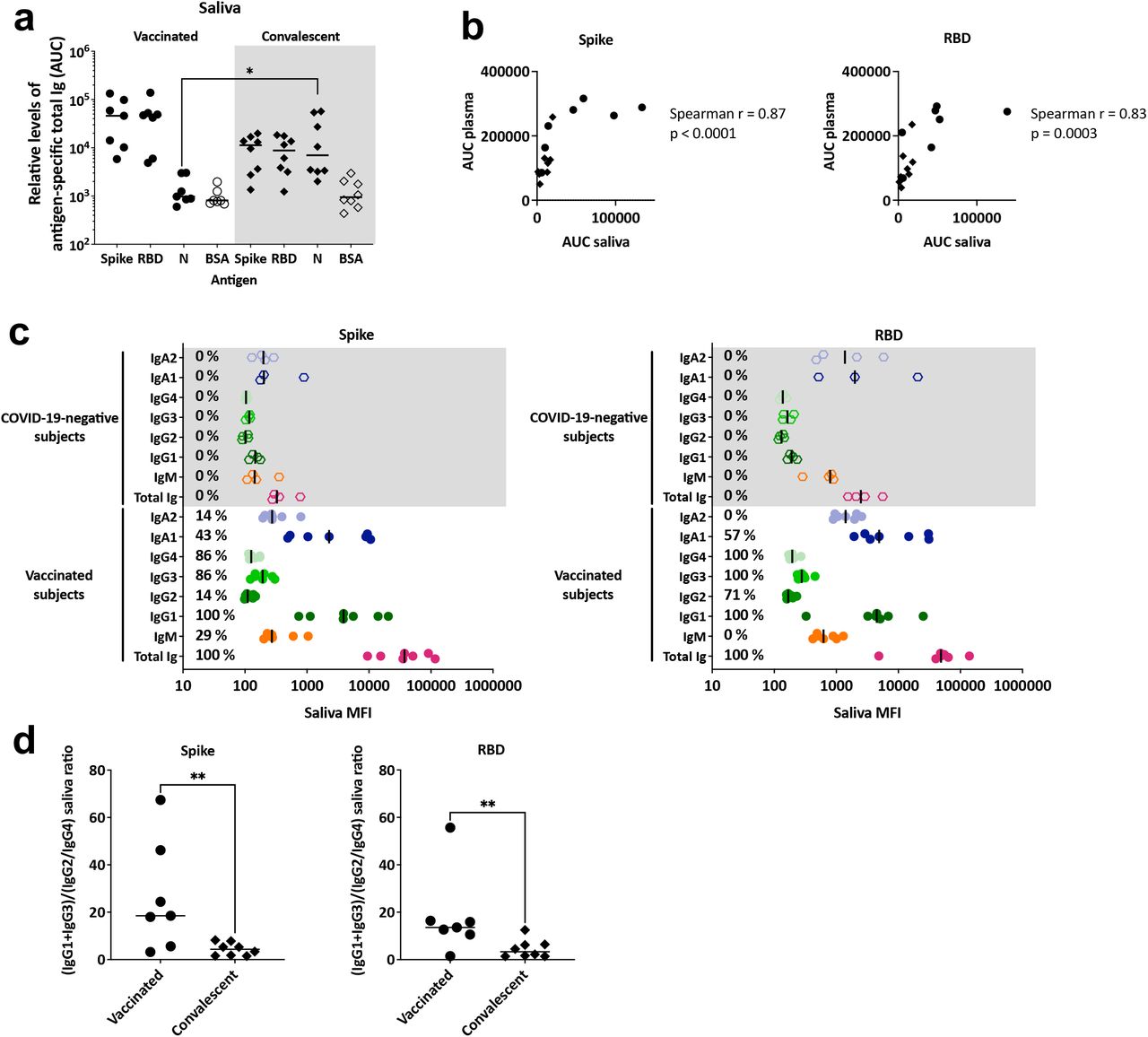
The saliva specimens were first titrated for total Ig levels against spike, RBD, S1, S2, and nucleoprotein antigens with BSA as a control (Fig. 7a, Supplemental Fig. 5a). All vaccinated subjects exhibited titrating amounts of saliva Ig against spike and S1, similar to those detected in plasma (Fig. 3b), although the saliva median effective concentration (EC50) titers of spike-specific Ig were on average 229-fold lower than the plasma titers. However, in contrast to 100% reactivity detected in plasma, only 5/7 saliva specimens from the vaccinated donors exhibited Ig reactivity against RBD and 2/7 against S2. No reactivity was detected against nucleoprotein or BSA, similar to what was seen with plasma specimens. In contrast, we detected Abs against spike, RBD, and nucleoprotein in saliva samples from convalescent COVID-19 patients (Fig. 7a). The presence of nucleoprotein-specific Abs in the saliva samples again differentiated convalescent from vaccinated subjects, similar to that seen in plasma. The AUCs of spike- and RBD-specific total Ig in the saliva of vaccinated and convalescent subjects did not differ significantly, although the median AUC of the vaccinated group was ∼3-fold higher than that of the convalescent group. Overall, a strong correlation was observed between spike- and RBD-specific total Ig levels in saliva versus plasma (Fig. 7b).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(a) Relative levels of plasma total Ig against spike, RBD, nucleocapsid, and BSA, as measured by area under the curves, (AUC), with specimens from seven vaccinated donors and eight COVID-19 convalescent patients tested in parallel. Statistical significance was calculated using a Kruskal-Wallis test. All data were generated using a multiplex bead antibody binding assay and plotted as mean fluorescent intensity (MFI). (b) Spearman correlation between spike- or RBD-specific total Ig AUC of plasma specimens versus saliva specimens from vaccinated (circles) and convalescent (diamonds) individuals. (c) Total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1 and IgA2 levels against spike and RBD in saliva specimens from seven vaccinated and four COVID-19 negative individuals. The black line represents the median for each group. The percentages of responders above the cut-off for each spike- or RBD-specific Ig isotype are indicated. (d) Spike- or RBD-specific ratios of IgG1+IgG3 MFIs over IgG2+IgG4 MFIs for vaccinated and convalescent saliva specimens. Statistical significance was calculated using a Mann-Whitney test.
The isotypes of spike- and RBD-specific Abs were subsequently examined in saliva at a 1:4 dilution. The COVID-19 mRNA vaccines induced anti-spike and anti-RBD IgG1 in the saliva of all vaccine recipients (Fig. 6c). A significant proportion of saliva samples also displayed IgG2-4 Abs against spike and RBD, even though the levels were lower than those of IgG1. Of the IgA isotypes in saliva, IgA1 responses against spike and RBD were more prevalent than their IgA2 counterparts. IgM responses were weak; only 2/7 saliva samples had detectable spike-specific IgM and none had RBD-specific IgM. The four control saliva specimens tested in parallel did not show any Ig isotype reactivity with spike or RBD.
Comparison of the plasma and saliva Ab profiles from the vaccinated versus convalescent groups is summarized in Fig. 3a,b. The plasma and saliva Ig isotype profiles were comparable to each other but differed between the vaccinated and convalescent group. The dominant isotype in saliva and plasma of vaccinated and convalescent groups was IgG1. However, the saliva IgG2-4 levels were higher in the vaccinated group, whereas the saliva IgM response was less prevalent in the vaccinated versus convalescent groups as seen in plasma (Supplemental Fig. 5a,b, [23]). Moreover, the saliva (IgG1+IgG3)/(IgG2+IgG4) ratios were higher in the vaccinated versus convalescent groups (p<0.01) (Fig. 7d), similar to that seen in plasma (Fig. 2c).
Discussion
This study provides the first evidence that COVID-19 mRNA vaccines induced plasma and saliva Ab responses dominated by IgG1. As compared to natural infection, vaccination also induced Ab responses marked by higher levels of IgG2-4 and greater ratios of (IgG1+IgG3)/(IgG2+IgG4). Vaccine-induced plasma Abs also displayed stronger capacity to mediate complement binding and activation. The differential potencies were apparent when the Ab levels were taken into account and when comparison was made with infection-induced Abs from earlier or later time points. By contrast, neutralization and ADCP potencies of vaccine- and infection-induced plasma Abs were comparable. The mRNA vaccine-induced Ab responses were also reported in a recent work with >8000 finger stick blood specimens to have higher seroconversion rates and greater cross-reactivity with SARS-CoV-1 and Middle Eastern respiratory syndrome (MERS)-CoV RBDs [29], further indicating the superior quality of vaccine-induced Ab responses.
We showed here that complement binding to spike-Ab immune complexes was stronger in plasma from vaccinated versus convalescent groups. However, the individual levels were highly variable for both groups, and no correlation was observed between the complement fixation capacity and the (IgG1+IgG3)/(IgG2+IgG4) ratios. A parameter known to influence complement binding is Fc glycosylation, as the removal of terminal galactose from IgG Fc glycans has been shown to decrease C1q binding and downstream classical complement activation without affecting FcγR-mediated functions [30,31]. Similarly, the sialylation of IgG Fc domains has been demonstrated to impair complement-dependent cytotoxicity [32]. The Fc glycan compositions of vaccine- and infection-induced Abs are yet unknown. The importance of complement binding/activation for protection against SARS-CoV-2 also requires further investigation. It should be noted, however, that human neutralizing monoclonal antibodies against SARS-CoV-2 require a functional Fc region capable of binding complement and engaging FcγR for ADCP and ADCC, for optimal protection as therapy [33].
We noted that while all vaccinated and convalescent individuals displayed plasma neutralization and ADCP activities, greater individual variability was seen in the levels of neutralization versus ADCP in both vaccinated and convalescent groups. Indeed, no correlation was observed between neutralization and ADCP activities, suggesting that these two functions are mediated by distinct Ig populations that may target different epitopes. We did not examine binding and functional Ab activities against the SARS-CoV-2 variants of concern such as B.1.1.7, B.1.351 and P.1. Most COVID-19 vaccines elicit Ab responses with weaker neutralization activities against these variants [34,35], but the effects of these and other emerging mutations on the non-neutralizing Fc-dependent functions are yet to be determined.
This study demonstrated that all vaccinated and convalescent individuals evaluated here had detectable saliva and plasma spike-specific Abs. Plasma and saliva anti-spike Ig levels after natural infection or vaccination correlated strongly. In a previous report, Abs against spike and nucleoprotein were detected similarly in plasma and saliva following SARS-CoV-2 infection [22]. These data indicate the potential use of saliva for monitoring of anti-spike Ab responses in vaccinated and convalescent individuals. However, lower positive responses were detected in saliva against RBD, S1, and S2 as compared to the intact spike antigen.
In conclusion, we demonstrate that, compared to natural infection, COVID-19 mRNA vaccines induce spike-specific Abs of diverse Ig isotypes and with potent complement binding capacities. The data highlight the superiority of vaccination in eliciting functional Ab responses against SARS-CoV-2.
Data Availability
Raw data are available upon written request to the corresponding author.
Author contributions
J.K. and C.E.H. wrote the manuscript. G.L., S.Z-P edited the manuscript. J.K., G.L., S.Z-P, C.U., and C.E.H. designed the experiments. J.K., G.L., V.I., and X.L. performed the experiments and collected the data. J.K., G.L., S.Z-P., C.U., and C.E.H. analyzed the data. K.Y.O., F.A., and B.L. provided protocols, antigens, cells and pseudovirus stocks. G.E-A., J.C.B., and S.L. obtained specimens. All authors read and approved the final manuscript.
Acknowledgments
We thank Dr. Florian Krammer for providing spike and RBD antigens, and all the donors for their contribution to the research. We would like to thank the expertise and assistance of Dr. Christopher Bare and the Dean’s Flow Cytometry CORE at Mount Sinai.
Footnotes
The authors do not have a commercial or other association that might post a conflict of interest.
This work was supported in part by the Department of Medicine of the Icahn School of Medicine at Mount Sinai Department of Medicine (to S.Z-P., C.E.H.); the Department of Microbiology and the Ward- Coleman estate for endowing the Ward-Coleman Chairs at the Icahn School of Medicine at Mount Sinai (to B.L.), the Department of Veterans Affairs [Merit Review Grant I01BX003860] (to C.E.H.) and [Research Career Scientist Award 1IK6BX004607] (to C.E.H.); the National Institutes of Health [grant AI139290] to C.E.H., [grants R01 AI123449, R21 AI1498033] to B.L, [grant R01 AI140909] to C.U. G.E-A and K.Y.O. were supported by Viral-Host Pathogenesis Training Grant T32 AI07647. K.Y.O was also supported by Training Grant F31 AI154739.
This manuscript has been deposited to medRxiv.
References



