
Abstract
Background Bacterial vaginosis is characterized by an imbalance of the vaginal microbiome in which the normally predominant lactobacilli are replaced by other bacterial species. Initiated by strains of the bacterium Gardnerella, a characteristic biofilm forms on the vaginal epithelium, explaining the typical presence of clue cells. This biofilm contributes to the resilience of the bacteria to antibiotic treatment, which may explain the frequent recurrence of BV.
Objective In this study, we investigate whether a therapy based on bacteriophage endolysins which specifically lyse Gardnerella, in particular the drug candidate PM-477, might be a promising alternative to broad-spectrum antibiotics and antiseptics.
Study design To identify Gardnerella specific endolysins, we searched for endolysin-encoding sequences in regions of Gardnerella genomes that are of prophage origin. This search identified fourteen homologous genes predicted to encode 1,4-beta-N-acetylmuramidase-type endolysins. When expressed in Escherichia coli and purified, the recombinant proteins demonstrated strong bactericidal activity against four different Gardnerella species. By shuffling the N-terminal catalytic domains and C-terminal cell wall-binding domains between the homologues, we produced 81 chimeric endolysins. These endolysins were tested for their activity and specificity in vitro and ex-vivo on vaginal samples from fifteen BV positive patients. Fluorescence in situ hybridization was used for visualization.
Results Several engineered endolysins were 10-fold more active than the most active wild-type enzymes. When tested against a panel of 20 Gardnerella strains, the most active endolysin, called PM-477, showed minimum inhibitory concentrations of 0.13–8 µg/ml. PM-477 had no effect on Lactobacillus strains or other species of vaginal bacteria. Furthermore, the efficacy of PM-477 was tested on vaginal samples from fifteen patients with either first time or recurring bacterial vaginosis. In fourteen cases, PM-477 killed the Gardnerella bacteria and physically dissolved the biofilms without affecting the remaining vaginal microbiome.
Conclusion The high selectivity and effectiveness in eliminating Gardnerella, both in cultures of isolated strains as well as in clinically derived samples of natural polymicrobial biofilms, makes PM-477 a promising drug candidate and an alternative to antibiotics for the treatment of bacterial vaginosis, especially in patients with frequent recurrence.
Background
Bacterial vaginosis (BV) is a very common disorder in women of reproductive age, with a prevalence estimated at 10–30 % worldwide 1,2. It is caused by an imbalance in the normal, healthy microbiome of the vagina that results mostly in discharge, odor and irritation. BV is associated with an increased risk of preterm delivery and low birthweight 3,4, infertility and early spontaneous abortion 5–7, and it is also a high risk factor for contracting sexually transmitted diseases, including HIV 8,9. Besides these potentially severe physiological consequences, frequently recurring and strongly symptomatic BV can have a huge negative impact on some women’s quality of life and psychological wellbeing.
The healthy vaginal microbiome is characterized by a low diversity and uniform colonization by a few species of Lactobacillus 10. In BV, the imbalance in the vaginal microbiome is represented predominantly by the loss of beneficial lactobacilli and overgrowth of bacteria such as Gardnerella, Prevotella, Atopobium, and Sneathia 11 and Lactobacillus iners 12. The current model of the etiology of BV focuses on the importance of Gardnerella species. It postulates that virulent strains of Gardnerella form an adherent biofilm on the vaginal epithelium in which other species can proliferate, resulting in a polymicrobial biofilm 13–17. This adherent biofilm is only weakly affected by the innate immune response 18, and the interplay with the host immune system is only partially understood 19. BV development may be triggered by sexual transmission of a mix of bacteria including Gardnerella, which can adhere to epithelial cells in the presence of lactobacilli. It has been suggested that, unlike strictly anaerobic bacteria associated with BV, Gardnerella, a facultative anaerobe, can tolerate the high redox potential created by the Lactobacillus-dominated healthy vaginal microbiome 15,20,21. The metabolism of Gardnerella spp., in turn, results in a local increase in pH and decrease of the redox potential, favoring growth of iron-dependent anaerobes 10 and suppression of lactobacilli 22. Recently, the species previously known as Gardnerella vaginalis was shown to comprise at least 13 different species of which four were named 23. Clinical studies indicated that various Gardnerella species may contribute differently to the pathogenesis of BV. A higher abundance of G. vaginalis and G. swidsinskii was found to be related with vaginal symptoms of abnormal odor and discharge and the relative abundances of G. vaginalis, G. swidsinskii, and G. piotii but not G. leopoldii were strongly associated with BV microbiome 24,25.
The therapies used currently to treat BV are broad-spectrum antibiotics, mainly metronidazole (MDZ), tinidazole (TDZ), or clindamycin (CLI), antiseptics (such as Octenisept®, based on the detergent-like octenidin), probiotics (preparations of lactobacilli), and prebiotics (lactate gels). Although all of these treatments may have some beneficial effects, none has proven to have satisfactory efficacy in preventing recurrence. Antibiotics are the most effective in quickly reducing the symptoms, but they are associated with a recurrence rate of up to 60 % within six months of treatment 26. One reason may be the presence of an antibiotic-tolerant biofilm in BV that shelters a reservoir of persister cells 26,27. Antibiotic treatment not only often fails to eradicate the biofilm, but can also lead to further dysbiosis of the vaginal microbiome and can promote candidiasis 28. Moreover, CLI can trigger pseudomebrannous colitis, cause antibiotics associated diarrhea and other gastro-intestinal side effects 29,30. Also, Gardnerella strains frequently develop resistance to MDZ and TDZ 31. Antiseptic treatments for BV have been less well studied than antibiotics but they are similar with regard to their effectiveness and are also associated with high recurrence rates and deterioration of the healthy vaginal microbiome 16,32. Studies of probiotics, and prebiotics are generally inconclusive: they appear not to result in lasting benefits 26. Vaginal microbiome transplantation after a course of antibiotic treatment had a promising effect in one study of five patients, but the approach is very resource-intense and hardly scalable 33. In conclusion, novel treatments that offer an alternative to antibiotics or that can be used in combination with antibiotics, are urgently needed.
Bacteriophage-encoded peptidoglycan hydrolases – also called endolysins or ‘enzybiotics’ – are a promising alternative to antibiotics 34,35. Produced towards the end of the lytic cycle in phage-infected bacteria, these enzymes cleave peptidoglycan in the bacterial cell wall, thus lysing the cells and releasing the progeny phages. They have been shown to be particularly effective against Gram-positive bacteria, which lack the Gram-negative outer membrane that might limit accessibility to the cell wall peptidoglycan. Endolysins have several advantages over antibiotics; not least, their very narrow host spectrum, which is usually limited to a single genus or even a single species 36, and their low propensity to generate resistance in their hosts37. Bacteriophages that invade Gram-positive bacteria encode a variety of highly diverse endolysins. In general, they have a modular structure consisting of one or more enzymatically active domain (EAD)s connected by a flexible interdomain linker to at least one cell wall-binding domain (CBD), typically located at the C-terminus of the protein. Both domains can contribute to the specificity for a given genus or species of bacteria 38.
Although no bacteriophages that infect Gardnerella species have yet been isolated, prophages encoding for endolysins are present in the Gardnerella genomes. Phage endolysins are a potentially novel and promising approach to treat BV because Gardnerella, now generally acknowledged as the central agent in the etiology of BV, has a thin cell wall. To develop endolysins specific for Gardnerella, we used an in silico approach to identify the genes encoding endolysins. We cloned and sequenced 14 such genes, demonstrated the lytic activity and specificity of purified recombinant proteins for Gardnerella spp. and engineered an enzyme with enhanced activity by domain shuffling. Moreover, we demonstrate the efficacy and specificity of this engineered endolysin on biofilms of Gardnerella in samples from patients with BV.
Material and Methods
Bacterial strains and culture conditions
Atopobium vaginae, Gardnerella spp., Lactobacillus iners, Mobiluncus spp, Streptococcus agalactiae, Prevotella bivia, and and were obtained from the Laboratory of Bacteriology, University of Ghent, Belgium. Other Lactobacillus spp. were obtained from the German Collection of Microorganisms (DSMZ). Gardnerella strains were either grown on chocolate (Choc) agar plates (Becton Dickinson) under anaerobic conditions in an anaerobic chamber equipped with anaerobic atmosphere generation bags (Sigma Aldrich) for 24–48 h. Lactobacillus spp. were cultured on Schaedler plates supplemented with vitamin K1 and 5 % sheep blood (Becton Dickinson), anaerobically at 37 °C for 24–48 h. A. vaginae, M. curtisii, M. mulieris, S. agalactiae, and P. bivia were either grown on Columbia blood agar plates, supplemented with 5 % sheep blood (Becton Dickinson) or Schaedler agar plates supplemented with vitamin K1 and 5 % sheep blood, anaerobically at 37 °C for 48 h. Escherichia coli BL21(DE3) cells (New England Biolabs) were used for protein expression and were grown on Luria Bertani (LB) agar plates (Becton Dickinson) or in LB (Luria/Miller) broth (Carl Roth) supplemented with appropriate selection antibiotic, aerobically at 37 °C. Liquid cultures of all species were done in New York City (NYC) broth III (10 mM HEPES (Sigma Aldrich), 15 g/L Proteose Peptone (Sigma Aldrich), 3.8 g/L yeast extract (Thermo Fisher Scientific), 86 mM sodium chloride (Carl Roth), 28 mM α-D-glucose (Sigma Aldrich)), supplemented with 10 % horse serum (HS) (Thermo Fisher Scientific).
Identification of Gardnerella genes encoding endolysin within prophage regions
Prophage regions in Gardnerella genomic sequences were identified by using PHASTER (https://phaster.ca/) to scan translated nucleotide sequences of Gardnerella genome entries in the NCBI database (https://blast.ncbi.nlm.nih.gov). Protein domain searches for endolysins were performed using INTERPRO (https://www.ebi.ac.uk/interpro/) to identify predicted enzymatically active domains in the corresponding amino acid sequence for each identified phage endolysin.
Gene cloning and overexpression of phage endolysins
Briefly, DNA of genes encoding wild-type or genetically engineered endolysins fused to a His6-tag was synthesized, cloned into the expression vector pET29b(+) and expressed in E. coli BL21(DE3). Protein expression was induced by TB (Terrific Broth Medium; 24 g/L Bacto Yeast Extract (Becton Dickinson), 12 g/L Bacto Tryptone (Becton Dickinson), 0.4 % glycerol (87%; Applichem), 0.02 mM potassium phosphate monobasic (Sigma Aldrich), 0.07 mM di-potassium hydrogen phosphate (Merck) and 1.5 % α-lactose monohydrate (Carl Roth)), by incubating at 25 °C for 24 h, with shaking at 250 rpm/min. The overexpressed proteins were purified by affinity chromatography on a nickel–nitrilotriacetic acid (Ni–NTA) affinity matrix (HISTrap column) eluted with 50 mM MES (Carl Roth) pH 7, 150 mM NaCl (Carl Roth), 250 mM imidazole (Carl Roth) and followed by size exclusion chromatography. Fractions were pooled and dialyzed against MES buffer (50 mM MES pH 5.5, 100 mM NaCl, 8 mM MgSO4 (Sigma Aldrich)). Protein concentration was determined at OD 260/280 nm or by using the Pierce™ BCA (bicinchoninic acid) protein assay kit (Thermo Fisher Scientific).
Culture-based assessment of bactericidal activity
Bacterial suspensions (OD600 of 0.1, corresponding to approximately 107–108 CFU/ml) were prepared by scraping the cells from confluently grown agar plates and diluting them into NYCIII+ 10 % HS, pH 5.5. Reactions were performed in triplicate by mixing 10 µl of endolysin (200 µg/ml) with 90 µl bacterial suspension in the wells of a 96-well plate. Ten µl MES buffer without the endolysin was used as a control. The 96-well reaction plate was incubated anaerobically at 37 °C for 5 h. Tenfold dilution series (10−1 to 10−6) of the cell reaction mixtures were prepared in NYCB+ 10 % HS and 2 µl of each dilution were spotted onto Choc agar plates. After anaerobic incubation at 37 °C for 48 h, colonies were counted, CFU/ml calculated, and the log10 reduction compared to MES buffer treated control was determined.
MIC assessment
The minimum inhibitory concentration (MIC), which is a standard measure of the activity of antimicrobials, was determined according to the Clinical and Laboratory Standards Institute protocol (2018) Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria 39. Bacterial suspensions of 105 to 106 CFU/ml in NYCIII+ 10 % HS were treated with a 1:2 dilution series of either PM-477 (starting concentration 64 µg/ml) or the antibiotics metronidazole (Gatt-Koller), tinidazole (Sigma Aldrich), or clindamycin (Clindamycin hydrochloride, Sigma Aldrich), starting with a concentration of 128 µg/ml and twofold dilution down to 0.0625 µg/ml. Controls for growth in the absence of antimicrobials were also included. OD620 was recorded by microplate reader (Tecan) after incubation at 37 °C for 48 h or, for some fast-growing strains, after incubation for 24 h, and compared to t=0 values.
Phase contrast microscopy
Dense bacterial suspensions were prepared in NYCIII+ 10 % HS either as Gardnerella monocultures or as a co-cultures of Gardnerella and Lactobacillus strains at final OD600= 4 for each strain. Bacterial mixtures were centrifuged at 3,800 x g for 7 min at room temperature in a benchtop centrifuge (Heraeus Instruments, Biofuge Pico). Supernatants were removed and cell pellets were resuspended in 50 µl of PM-477 (460 µg/ml) or in MES buffer alone as a negative control and incubated for 5 h at 37 °C under anaerobic conditions. Subsequently, 5 µl of each bacterial suspension were placed on a glass slide, covered with a glass coverslip and observed by phase contrast microscopy (Olympus BX41 microscope, magnification 1000x).
For the comparison of bacteriolytic and bactericidal effects, bacterial suspensions of Gardnerella swidsinskii and Gardnerella vaginalis were prepared in NYCIII+ 10 % HS at a final OD600= 1. Bacterial suspensions of 50 µl were centrifuged at 3,800 x g for 7 min at room temperature in a benchtop centrifuge (Heraeus Instruments, Biofuge Pico). Supernatants were removed and pellets were resuspended in 50 µl of MES buffer containing PM-477 at 400 µg/ml, 200 µg/ml, 100 µg/ml, 50 µg/ml or 25 µg/ml, or in MES buffer alone as a negative control, and incubated for 5 h at 37 °C in anaerobic conditions. Subsequently, 5 µl of each bacterial suspension were observed by phase contrast microscopy as above. To determine viability, 10 µl of the reaction mixture was serially diluted and spotted onto Choc agar plates. After incubation at 37 °C for 48 h, the colonies were counted and CFU/ml was calculated.
Lytic effects of PM-477 and Octenisept® on ex vivo BV patient samples as detected by fluorescence in situ hybridization (FISH) microscopy
Vaginal samples were collected from randomly selected 24–49 year-old Caucasian women who were previously diagnosed with BV according to the Amsel criteria 40. In addition, the presence of clue cells (epithelial cells covered with cohesive Gardnerella bacteria) was determined by FISH microscopy 16. Fresh vaginal smears were collected with a swab and put in ESwab(tm) 493C02 pre-filled vials (COPAN Diagnostics). The vials were vortexed gently, resulting in a suspension that contained epithelial cells and vaginal bacteria. Four consecutive vaginal smears from each patient were pooled. The effects of 0.2, 2, 20 and 200 µg/ml of PM-477 after 2, 6, and 24 h, respectively, were tested either at room temperature or 36 °C. Since the most pronounced lytic effect on Gardnerella was observed after 24 h exposure to 200 µg/ml PM-477 at 36 °C (Supplementary Table 1), this condition was used for further studies. Subsequently, vaginal swabs from 15 BV patients were collected, aliquoted, and mixed 1:1 (v/v) with 400 µg/ml PM-477 (final concentration of 200 µg/ml) or with MES buffer as a negative control, or mixed 1:20 (v/v) with the disinfectant Octenisept® (Schülke) and incubated for 24 h at 36 °C. Untreated samples were also included in the analyses as baseline controls. The samples were fixed with Carnoy solution (alcohol/chloroform/acetic acid 6/3/1 by volume) 41. Fields of 10 mm x 10 mm were marked on SuperFrost slides (Langenbrinck, Emmendingen, Germany) with a PAP pen (Kisker-Biotech, Steinfurt, Germany). Five μl aliquots of vortexed, fixed vaginal smear sample were dropped onto the marked field. The slides were dried for 60 min at 50 °C before FISH analysis.
For multicolor FISH analyses, we used the Gard662 Gardnerella-specific DNA hybridization probe 42 and the universal bacterial probe Eub338 43. Oligonucleotide probes were synthesized with a fluorescent dye (Cy3 or Cy5). The hybridization was performed at 50 °C as previously described 41. The probe Ato291 was used for Atopobium, Liner23-2 for L. iners, and Lcrisp16-1 for L. crispatus 42. DAPI was used to visualize the vaginal epithelial cells (microphotographs not shown). Changes in density and distribution of microbial species were monitored by using a Nikon Eclipse 80i fluorescence microscope, a Nikon SHG1 camera and accompanying software (Nikon, Tokyo, Japan). To avoid dispersion related biases (the thickness of smears on the glass slide and the microbial distribution is uneven), bacteria were enumerated for representative areas containing 10 epithelial cells as follows. For bacteria in low densities (e.g. lactobacilli), larger areas, including at least 10 microscopic fields were evaluated and mean cell numbers were expressed in relation to 10 epithelial cells. In areas with high local bacterial concentrations (e.g. in patches of biofilms), where cells are adjacent and in part overlapping, each 10 µm x 10 µm area covered with bacteria was counted as 500 bacterial cells. The numbers were rounded to full 100s.
Statistical analysis
Where appropriate, data were log-normalized prior to applying statistical tests (e.g. for CFU/ml values, and as indicated in the figure legends). When only two groups were compared, the unpaired two-tailed Student’s t-test was used as indicated in the respective figure legends. Multiple groups were compared by two-tailed one-way ANOVA tests. The software used for statistical analyses was GraphPad Prism8. Differences between groups were considered statistically significant when P ≤ 0.05.
Results
Gardnerella genomes contain prophage genes that encode active endolysins
Although no bacteriophages of Gardnerella species are known, multiple Gardnerella genomes that have been sequenced contain DNA regions that are predicted to be of prophage origin 44,45. To identify endolysins that might target specifically Gardnerella, we used Basic Local Alignment Search Tool (BLAST) to search for sequences with similarities to known endolysin genes in the prophage regions of published Gardnerella genomes. A total of 14 genes were identified that encoded 1,4-beta-N-acetylmuramidases, with sequence similarity between 87 to 98 %, when compared to each other. Twelve genes encode a 306 residues protein whereas two encode similar proteins that lack 55 residues at the C-terminus (Supplementary Table 2). The sequences all encode domain structures common to all known endolysins: an N-terminal EAD of the glycoside hydrolase family 25 (GH25) and a CBD, comprising two CW_7 motifs, homologous to the CBD of Cpl-7 lysozyme, encoded by the Streptococcus pneumoniae bacteriophage Cp-7. The sequences of these putative endolysins are 90–94% identical to some proteins encoded by the genomes of Atopobium and Bifidobacteriaceae but < 70 % identical to other endolysins, which may already indicate a high specificity for the genus Gardnerella (data not shown).
We expressed the 14 predicted endolysin genes (EL1–EL14) as recombinant proteins in E. coli, and ten (EL 1–7 and EL 10–12) yielded sufficient protein to perform further experiments. Their lytic activities were tested on representatives of the four Gardnerella species: G. vaginalis (ATCC 14018T), G. leopoldii (UGent 09.48), G. piotii (UGent 18.01T), and G. swidsinskii (GS10234). Bacterial suspensions of 107-108 CFU/ml were incubated with each of the 10 endolysins (at 20 µg/ml) or with buffer as a negative control. After a 5 h incubation, we plated the suspensions and quantified the surviving bacteria, measured as colony-forming units (CFU)/ml (Figure 1). Most of the endolysins reduced the CFU/ml of all four Gardnerella species by multiple log10 units. Only EL5 and EL6 (one of the two truncated proteins mentioned above) were completely inactive in this assay. EL3 and EL10 were the most active against all four Gardnerella species tested, G. swidsinskii GS10234 being the most susceptible with a > 5 log10-fold reduction in CFU/ml (Figure 1).

Wild-type endolysins are highly active against four different Gardnerella species. Suspensions of G. vaginalis (ATCC 14018T), G. leopoldii (UGent 09.48), G. piotii (UGent 18.01T), and G. swidsinskii (GS10234) were treated with recombinant endolysins prepared by expression of the naturally occurring genes (EL1–EL7 and EL10–EL12) (see Methods) and the log10 reduction in viable CFU/ml was determined. The log10 reduction was calculated by comparing the CFU/ml after endolysin treatment with that of the buffer control (w/o). LOD indicates the limit of detection.
Engineering of a Gardnerella endolysin with enhanced bactericidal activity
To increase the lytic activity of the endolysins identified above, we used domain shuffling 46,47 to recombine systematically ten of the EADs encoded by the newly identified endolysin genes EL 1–7 and EL 10–12 (annotated as H1–7 and H10–12) with nine CBDs encoded by EL1–5, EL7 and EL10–12 (annotated as B1–5, B7, and B10–12; since EL6 was inactive, we did not use its truncated CBD). All of the 81 resulting chimeric proteins were expressed in E. coli and purified by means of their N-terminal His tags. We tested these chimeric endolysins for their bactericidal activity on the abovementioned Gardnerella strains and compared their activity to that of the 10 wild-type endolysins (Figure 2).

Domain swapping increases the lytic activity of engineered endolysins. Eighty-one constructs comprising all possible combinations of the EAD (H) and CBD (B) domains of 10 wild-type endolysins were tested on four different Gardnerella species as in Figure 1. The values represent the log10 of the ratio of viable cells (CFU/ml; mean of triplicate assays) in endolysin-treated suspensions of Gardnerella compared to buffer control-treated suspensions of the species, as indicated. Blue shades indicate higher bactericidal effect (high reduction in viable CFU/ml) compared to the wild-type endolysin, red shades indicate lower bactericidal effect, and white coloring indicates equal reduction compared to wild-type endolysin (boxed in black). The endolysin candidate with the highest average reduction over four Gardnerella species is boxed in yellow.
Overall, G. swidsinskii GS10234 was most sensitive to most endolysins – wild-type and chimeric. Two of the chimeric endolysins, H2B10 and H2B11, were more active than any wild-type enzyme against all four Gardnerella strains, whereas others were more active than any wild-type endolysin against some strains (e.g. H2B12, H5B3, H5B4, H7B3 and H7B11) (Figure 2). Overall, chimeric endolysins combining domains H2, H7 or H10 with domains B10, B11 or B12 showed highest bactericidal activity. The combination of H2 and B10 was most effective against any of the four strains and reduced their viability by an average of 4.3 log10 CFU/ml. We designated this construct PM-477 and chose it for further development as a candidate for the treatment of BV.
Specificity of PM-477 for the genus Gardnerella
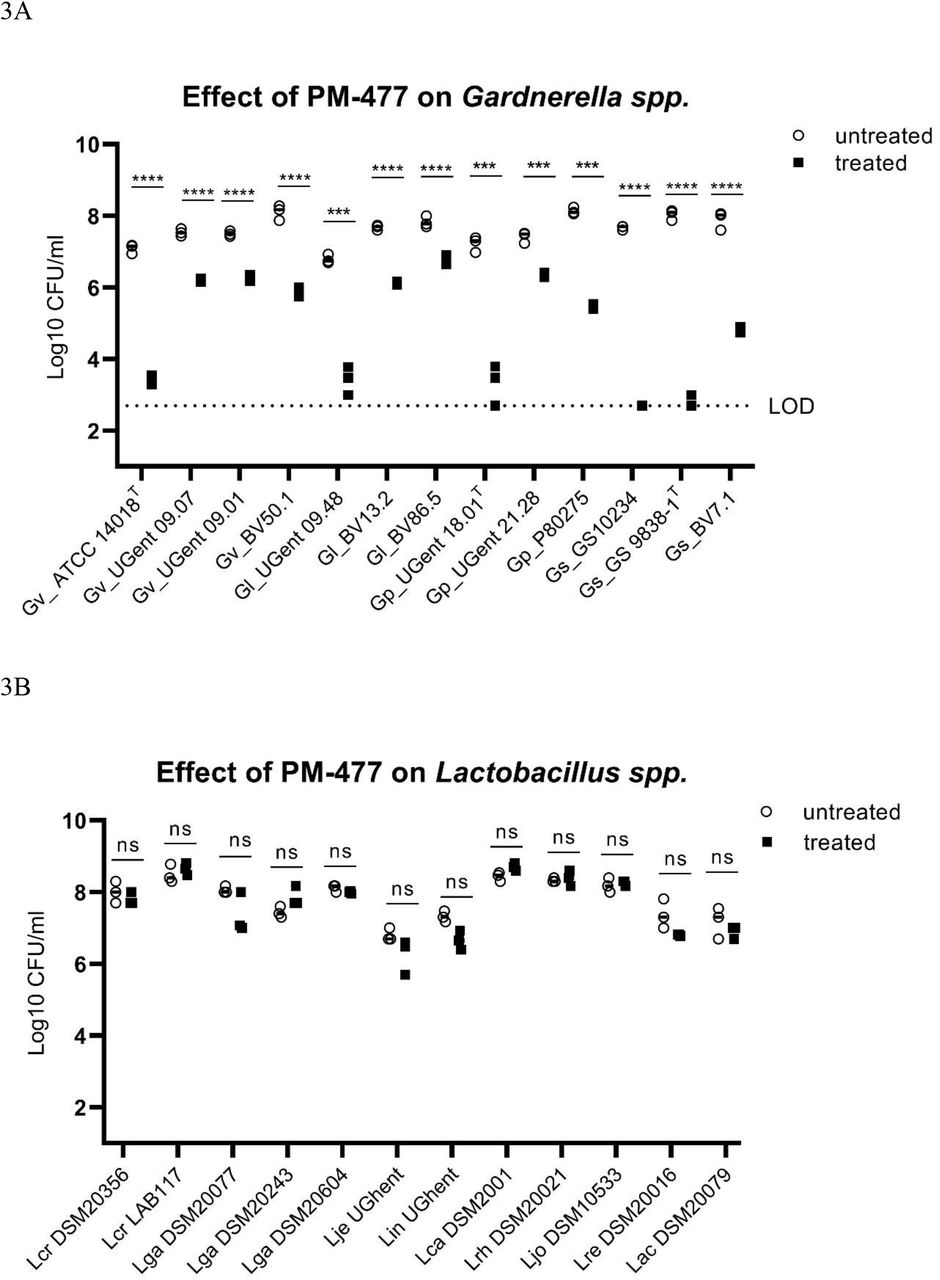
We characterized the activity and specificity of PM-477 against panels of 13 Gardnerella strains of the four species described above, 12 strains of vaginal lactobacilli (including L. crispatus, L. jensenii and L. gasseri, selected as representative of the healthy vaginal microbiome) and nine other species typically found in BV, by using the quantitative assay for bacterial viability described above. The engineered endolysin caused a 4–5 log10-fold loss of viability in three Gardnerella strains (Gs_GS10234, Gs_GS9838-1T, Gp_UGent 18.01T), a 2– 3 log10-fold loss of viability in five strains (Gv_ATCC 14018T, Gv_BV50.1, Gl_UGent 09.48, Gp_P80275, Gs_BV7.1), and a 1–2 log10-fold loss of viability in a further five strains (Gv_UGent 09.07, Gv_UGent 09.01, Gl_BV13.2, Gl_BV86.5, Gp_UGent 21.28) (Figure 3A).

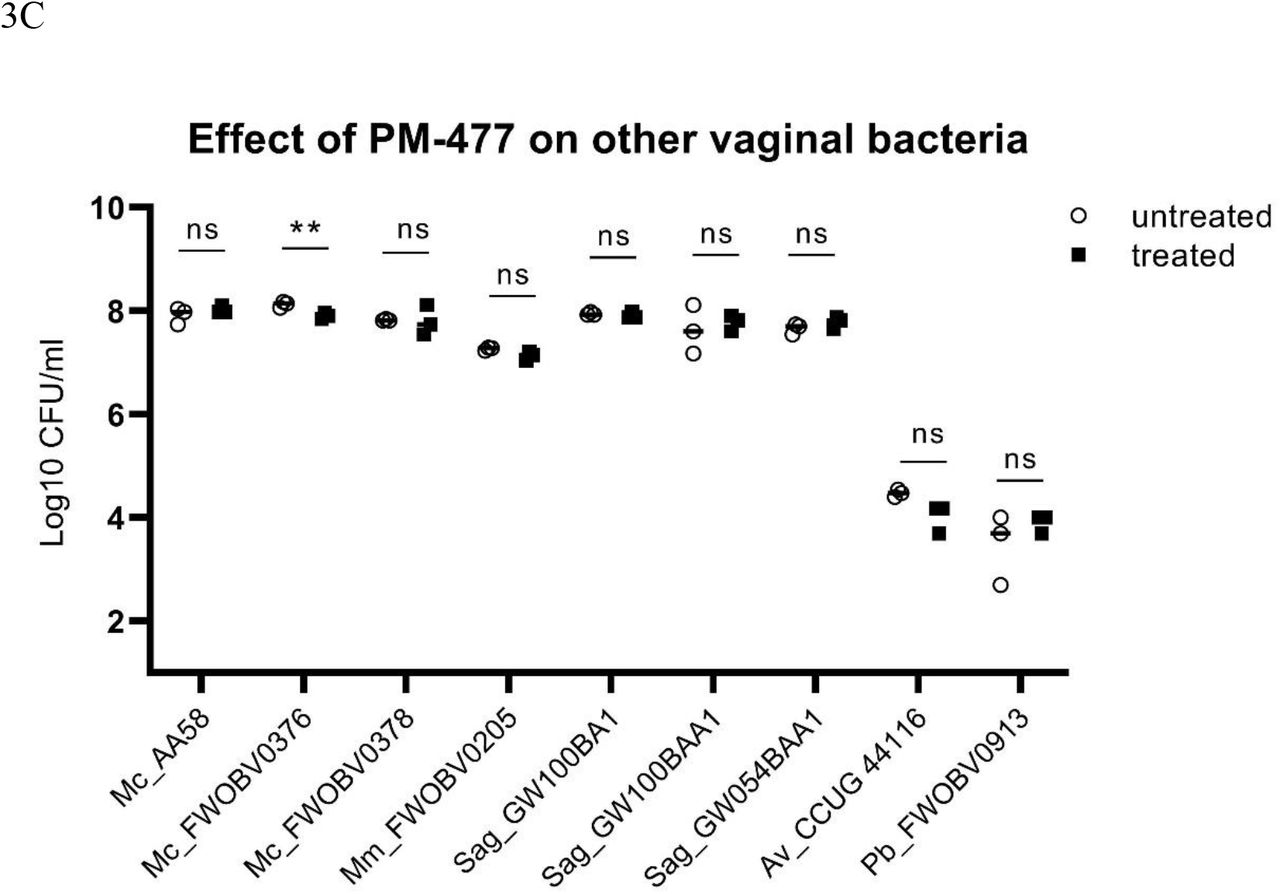
PM-477 is highly specific for Gardnerella and spares lactobacilli and other BV associated microbiome components. (a) Gardnerella strains, (b) Lactobacillus spp., (c) and Mobiluncus curtisii, Mobiluncus mulieris, Streptococcus agalactiae, Atopobium vaginae and Prevotella bivia were tested for their susceptibility to PM-477 (20 µg/ml for 5h; treated, black squares) and compared to buffer control-treated (untreated, white circles) bacteria suspensions. For statistical purposes CFU/ml values from the untreated and the treated groups were log-normalized prior to applying the unpaired two-tailed multiple Student’s t-test. P-values ≤ 0.05 were considered statistically significant; ** P≤0.01,***P≤0.001,**** P≤0.0001.
The same treatment of lactobacilli, by contrast, had no statistically significant effect on the viability of any of the 12 strains tested (Figure 3B). It also had no effect on strains of Atopobium vaginae, Mobiluncus mulieris, Prevotella bivia and Streptococcus agalactiae, and had only a minor effect one of three strains tested of Mobiluncus curtisii (Figure 3C). (In the cases of Atopobium vaginae and Prevotella bivia the cells either grew poorly or died rapidly upon transfer from agar plates to suspension, even without treatment.) Thus, the engineered endolysin PM-477 is highly selective for Gardnerella: it kills strains of each of the four main species 23, without harming beneficial lactobacilli or other species typical of the vaginal microbiome.
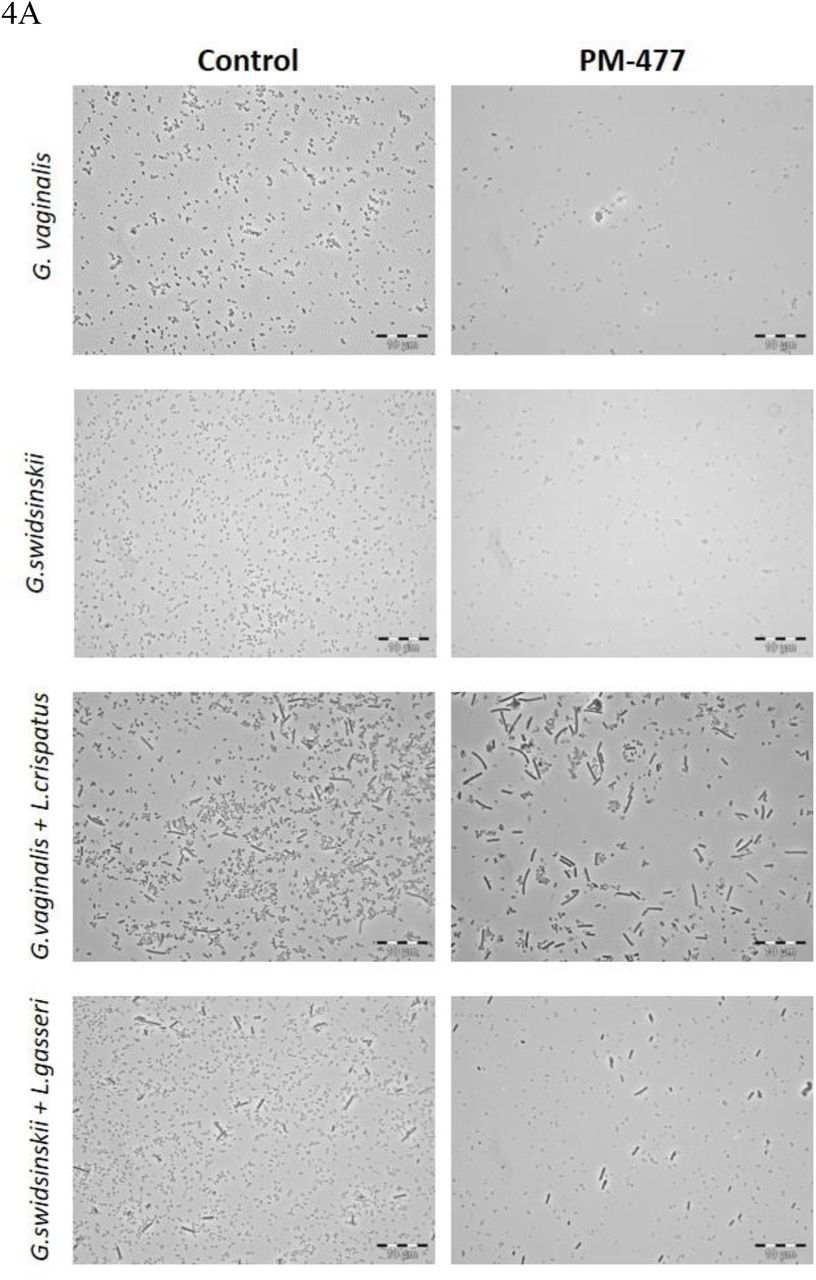
The genus-specific bacteriolytic effect of PM-477 was confirmed by microscopy in mixed cultures of Gardnerella and lactobacilli. These two genera can be distinguished by their distinct morphologies: Gardnerella cells are small coccoidal rods, whereas lactobacilli form mostly long and thick rods. PM-477 (at 460 µg/ml for 5 h) lysed G. vaginalis and G. swidsinskii cells in mono-culture and selectively lysed them in mixed cultures of G. vaginalis and L. crispatus and of G. swidsinskii and L. gasseri, respectively, while the lactobacilli in these mixed cultures were unaffected (Figure 4A).



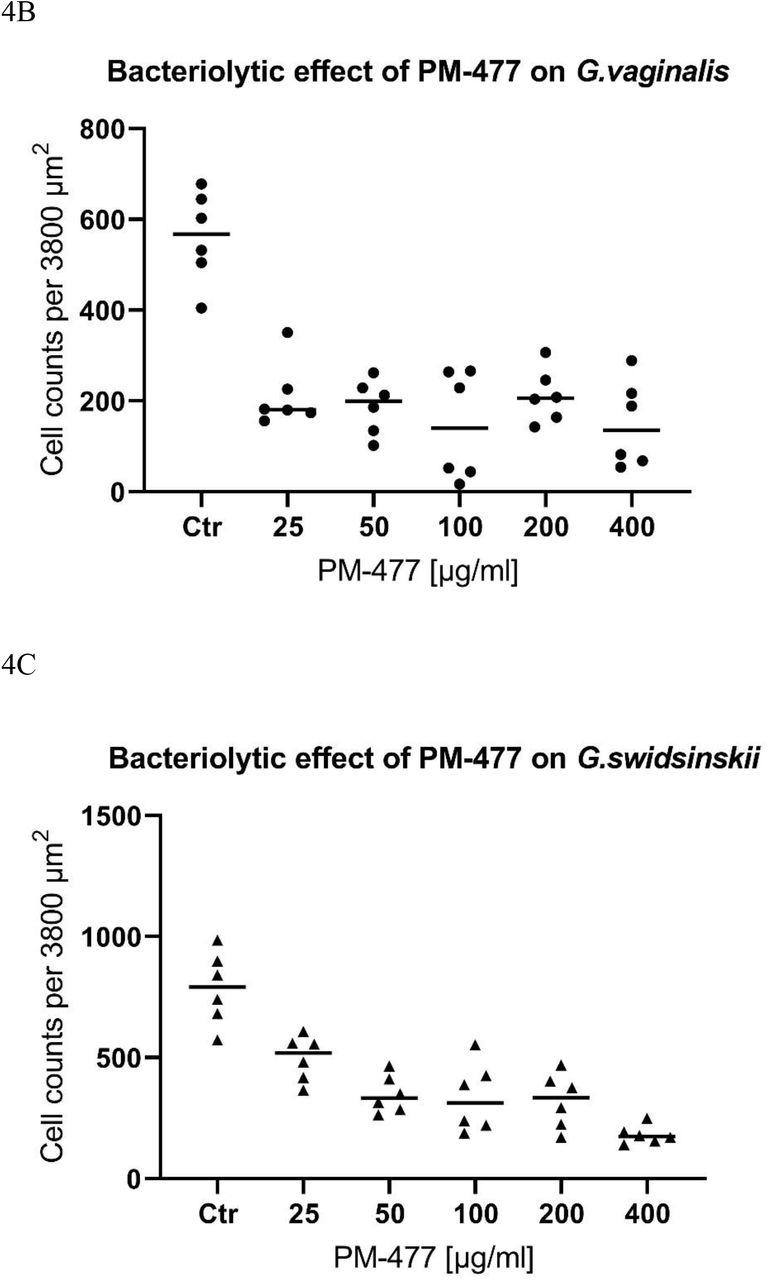
PM-477 selectively kills Gardnerella in a co-culture with lactobacilli. A, Phase contrast microscopy of dense cultures of G. vaginalis (ATCC14018T) (first row) and G. swidsinskii (GS10234) (second row), or co-cultures of G. vaginalis (ATCC14018T) and L. crispatus (LABCRI FB020-08c) (third row), as well as G. swidsinskii (GS10234) and L. gasseri (LAB GAS LMG 9203T) (fourth row) treated with PM-477 (460 µg/ml for 5h) or with buffer (control). B and C, Cell counts of Gardnerella cells determined by phase contrast microscopy after treatment with 25–400 µg/ml PM-477 for 5h or with buffer (Ctr). D and E, Viable cells remaining (log10 reduction CFU/ml compared to the buffer-treated control) in suspensions of Gardnerella treated with 25–400 µg/ml PM-477 for 5h.
We compared the bactericidal and bacteriolytic effects of PM-477 on G. vaginalis and G. swidsinskii by treating suspensions of the bacteria with 25–400 µg/ml PM-477 for 5 h and then counting the cell numbers by microscopy and determining their viability by quantitative plating (Figure 4B–E). The highest concentration of PM-477 (400 µg/ml) decreased the number of cell-like structures by only 80–90 % (Figure 4B, C), whereas the same treatment resulted in a ≥ 5.6 log10 reduction in viable cells. This indicates that PM-477 may kill before fully dissolving bacterial cells, resulting in visible but dead cell-like structures seen under the microscope (Figure 4D, E).
Efficacy of PM 477 compared to standard antimicrobials
To compare the efficacy of PM-477 in suspension to the standard antimicrobials used to treat BV, we determined the minimum inhibitory concentration (MIC)s of PM-477, CLI, MDZ and TDZ for 20 strains of Gardnerella by following the Clinical and Laboratory Standards Institute protocol 39. All the Gardnerella species tested were highly susceptible to PM-477 with a MIC90 value (the MIC value which inhibits 90 % of strains) of 8 µg/ml (Table 1).
Minimum inhibitory concentration (MIC) of PM-477 and the antibiotics CLI, MDZ, and TDZ for various Gardnerella spp. and Lactobacillus spp. (R) Resistance, defined as ≥ 32 µg/ml for MDZ and TDZ, and ≥ 8 µg/ml for CLI, according to international standards.
The G. swidsinskii strains had MICs in the range of 0.25–1 µg/ml, whereas the G. vaginalis strains had MICs of 0.13–8 µg/ml, and G. leopoldii and G. piotii strains had MICs of 1–8 µg/ml. Among this small number of strains tested for each species, there was no evidence that the four Gardnerella species differed with regard to their susceptibility to PM-477. Whereas 12 of the 20 Gardnerella strains were resistant to MDZ and 15 were resistant TDZ, CLI was effective against all strains at MICs ≤ 1 µg/ml. All tested strains of three Lactobacillus species (L. crispatus, L. gasseri, and L. jensenii) were resistant to the highest concentrations of PM-477, MDZ, and TDZ (MICs > 128 µg/ml), whereas only one strain was resistant to CLI.
Efficacy of PM-477 against biofilms of Gardnerella in human BV samples
To analyze the efficacy of PM-477 in a physiological environment that resembles closely the in vivo situation, we treated vaginal swabs from 15 BV patients, and analyzed them by fluorescence in-situ hybridization (FISH).
Areas of strong yellow fluorescence in the untreated (baseline) and buffer control-treated samples indicated the presence of Gardnerella-dominated biofilm on exfoliated vaginal epithelial cells (visible due to their pale autofluorescence and clearly corresponding to DAPI counterstain), called clue cells. At baseline and after buffer treatment, the yellow fluorescence of the Gardnerella probe outshines the red fluorescence of the probe for all bacteria, indicating that the biofilm is composed in large part of Gardnerella cells. Figure 5A shows a total resolution of the dense yellow staining on the epithelial cells upon treatment with PM-477. Therefore, PM 477 proved strong bacteriolytic activity also in this ex-vivo setting. The specificity of PM-477 was demonstrated by the remaining red signal from the universal bacterial probe, indicating mostly non-Gardnerella cells. Only very few individual Gardnerella cells seemed to remain associated with or in between the epithelial cells. However, the comparison of bacteriolytic and bactericidal effect in Figure 4B-E would indicate that these might be dead cell-like structures with sufficient DNA associated to them to hybridize the FISH probes. In contrast to PM-477, Octenisept killed and lysed both the Gardnerella and other bacterial cells non-specifically, as indicated by the visible reduction of both types of fluorescence.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
PM-477 dissolves the Gardnerella-dominated biofilm on exfoliated vaginal epithelial cells from BV patients. A, Representative examples of multi-color FISH with Gard Cy3 probe (Gardnerella, yellow fluorescence) and universal Eub 338 Cy5 probe (all bacteria, red fluorescence) in 400x magnification (where not stated otherwise) for patients 11, 12 and 15. Baseline (prior to treatment): Gardnerella bacteria are cohesive and adherent to epithelial cells forming dense biofilm patches on the surface of clue cells which is apparent due to background fluorescence. The attachment of Gardnerella biofilm to epithelial cells can be seen especially well in DAPI stains and on zoomed original microphotographs (DAPI microphotographs are added to supplementary material together with all other original microphotographs of Figure 5) Control: Gardnerella dominated biofilms in buffer-treated controls after 24 h incubation are still dense in patient 11 and 12 and start to partially disintegrate in patient 15. PM-477: After incubation with PM-477, the Gardnerella biofilm is completely disintegrated in all patients. The disappearance of intense yellow fluorescent Gardnerella bacteria unmasks the red fluorescence of still intact bacterial mass. Octenisept: similar to PM-477, Octenisept massively reduces Gardnerella (yellow) but additionally nearly completely eliminates any other non-Gardnerella bacteria (red). B, Gardnerella cell density in samples of patients 1-15 expressed as the number of yellow bacteria counted in areas containing 10 epithelial cells (see methods for a detailed description of the counting method)., p-values were calculated with ANOVA in GraphPad Prism.
The effect of treatment was also quantified by counting the number of bacterial cells per 10 epithelial cells in representative fields of vision (Figure 5B). When compared with the baseline samples, no significant changes were observed upon incubation with the buffer control, indicating that the vaginal microbiome was generally stable over the time-course of the experiment. PM-477 significantly reduced Gardnerella cells in the patient samples (median of 300 cells/10 epithelial cells) when compared to baseline (median of 1200 cells/10 epithelial cells, p=0.02) and when compared to the buffer control-treated samples (median of 900 cells/ 10 epithelial cells, p=0.007): 3 of the 15 samples showed a >500-fold reduction in Gardnerella load, 4 showed a >100-fold reduction, 7 a >10-fold reduction and 11 a >2-fold reduction (data for individual patient samples are presented in Supplementary Table 3).
The effects of PM-477 on Gardnerella were similar to those of Octenisept®. Unlike PM-477, however, the lytic activity of the disinfectant was not specific for Gardnerella but affected all bacteria (Figure 5A, Supplementary Table 3).
Two other potential BV associated bacteria, A. vaginae and L. iners, as well as L. crispatus, were visualized by using selective FISH probes and their cell numbers per 10 epithelial cells were counted (Supplementary Table 3). A. vaginae, L. crispatus, and L. iners were detected in 10, 3 and 8 out of the 15 samples, respectively. Treatment with PM-477 reduced the median cell counts of L. iners when compared to treatment with the buffer control in all samples whereas the numbers of L. crispatus and A. vaginae bacteria were unaffected. Octenisept® strongly reduced the cell counts of all three species.
In summary, these experiments provide a proof of the efficacy and selectivity of PM-477 for the genus Gardnerella in vitro and also in the native BV biofilm.
Discussion
This is the first study to show that prophage-derived endolysins active on Gardnerella could be used as an innovative treatment for bacterial vaginosis. Although no Gardnerella phages were ever isolated in a lab, we were able to identify putative endolysins in silico. We expressed them as recombinant proteins, improved their potency by systematic domain shuffling, demonstrated their efficacy and selectivity in vitro, and, ultimately, on the native polymicrobial biofilms in vaginal swabs from BV patients.
The currently available therapy of bacterial vaginosis is unsatisfactory. Antibiotic-centered ‘standard-of-care’ treatment for BV is associated with a recurrence rate of ∼60 % within 6 months, which may be explained by low efficacy of antibiotics on bacteria growing as biofilms, but also by selective destruction of beneficial lactobacilli, which are thereby prevented from re-establishing a protective environment 26. Bacteriophage-derived endolysins have been described as a potential alternative to antibiotics; they are both highly active on mucosal biofilms and selective for individual genera or even species of bacteria 35. Neither endolysins nor whole phages specific to Gardnerella have been isolated previously, although the existence of Gardnerella-specific phages has been postulated on the basis of genomic analyses 44. We identified Gardnerella-specific endolysin genes in the prophage-like genome sequences of strains of Gardnerella. A similar approach was reported earlier, in which the phage lysin PlyCD was identified by searching prophage-like sequences in the genomes of Clostridium difficile strains 48. The fact that it is possible to predict and generate endolysins from prophage encoded sequences in the genomes of unrelated species (C. difficile and Gardnerella) may indicate a wide application of this approach, which is easily adaptable to other pathogenic bacteria.
We could show that the wild-type endolysins we identified and, even more so, our engineered constructs, are highly bactericidal and selective for the genus Gardnerella.
Our engineered endolysin PM-477 is superior to the current standard of care antibiotics in that it is active in the low µg/ml range and effective against all 20 of the Gardnerella strains we tested, covering all four named species. In total, 60 % of the 20 strains were resistant to MDZ and 75 % to TZD, reinforcing the notion that PM-477 may be more effective against BV than currently used standard antibiotics.
The high resistance rate for MDZ, which was also reported elsewhere 31, does not support the finding that MDZ treatment temporarily relieves the symptoms in a majority of cases 26. However, long-term cure is mostly not achieved with MDZ indicating that their effectiveness may be reduced due to the high resistance rate of Gardnerella strains 49.
In our study, CLI was also effective against all the strains of different Gardnerella genotypes we tested. Curiously, however, CLI has not shown superior efficacy when compared to MDZ in clinical trials 26, which may indicate that BV biofilms are more tolerant to CLI than they are of MDZ, thus compensating the higher rate of resistance against MDZ.
In conclusion, PM-477 is superior to all tested antibiotics because, unlike nitroimidazoles, it is active on all Gardnerella strains and, unlike CLI, it does not kill lactobacilli. Antibiotics may have the advantage over PM-477 of being active against a broader spectrum of pathogens. But, unlike Gardnerella, these pathogens may not be pivotal in vaginal biofilm formation as is the case in BV. Also, antibiotics typically have low activity on bacteria in biofilms 27 whereas our ex-vivo data indicate that PM-477 works well on Gardnerella in vaginal biofilms.
Progress has been made in developing animal models for BV. For example, it was possible to model some of the clinical characteristics of BV, such as epithelial exfoliation and clue cells upon infection of mice with Gardnerella 50. However, it was not possible so far to model the increase in pH and the diverse microbiome composition of human BV in animals. Therefore, the next best and, arguably, more biologically relevant model to test the efficacy of PM-477 is to use ex-vivo vaginal samples from BV patients, which contain clue cells, i.e. exfoliated epithelial cells covered with a polymicrobial biofilm. The microbial populations of all the ex vivo samples that we collected from 15 individual BV patients were clearly dominated by Gardnerella and a substantial part of the Gardnerella cells adhered to the epithelial cells. Treatment with PM-477 fully resolved the dense staining due to Gardnerella cells on the epithelial cells while not affecting non-Gardnerella cells. Thus, we conclude that PM-477 quantitatively removes Gardnerella cells inside the polymicrobial biofilm on vaginal epithelial cells ex vivo. Furthermore, it is effective in the environment of the vaginal fluid despite the putative presence of proteases 51 and other substances that might reduce its activity.
PM-477 shows promising results in vitro and ex vivo but we do not know yet whether selectively removing Gardnerella will be sufficient to resolve the clinical symptoms of BV, which are caused by a polymicrobial biofilm, and to allow lactobacilli to re-colonize the vagina. The increasing evidence of the central role that Gardnerella plays in the etiology of BV 15 and the observation that BV seems not to be possible without Gardnerella 22, indicates that this might well be the case. The biomass of the polymicrobial biofilm consists largely of Gardnerella cells 52,53, so selective removal of Gardnerella should decimate the biofilm. Moreover, Gardnerella mediates adhesion to the epithelial cells, whereas other bacteria, especially in smaller numbers, may not be able to initiate biofilm formation on these cells 54. Other pathogens, especially Prevotella bivia, synergize with Gardnerella, so when the latter is removed the viability of the other pathogens is likely reduced as well 55,56. Removal of Gardnerella (and the concomitant reduction in the viability of P. bivia) would reduce the levels of sialidase produced by Gardnerella and P. bivia and allow the mucus layer to reform, in turn reducing the adherence of the remaining BV pathogens and depriving them of carbon sources 57. Finally, the active suppression of lactobacilli by Gardnerella 22 would be released, allowing recolonization by benign bacteria. This hypothesis has been investigated with in vitro biofilm models 17,20, and PM-477 as a selective antimicrobial agent may be a very useful tool to further investigate the role of Gardnerella in ex vivo as well as in in vivo studies.
In summary, we report here that an engineered, phage-derived endolysin has the potential to completely disrupt the Gardnerella-dominated biofilm in vaginal swabs from patients with BV. PM-477 is, thus a promising candidate for an effective treatment of recurrent BV, which remains, despite decades of research, a major public health concern.
Data Availability
All relevant data is disclosed in the manuscript.
Acknowledgements
We thank Grégory Resch and Life Science Editors for scientific editing assistance during the preparation of the manuscript.
Footnotes
Disclosure Conflicts of interests have been declared by the following authors: CL, LT, VO, are employees of PhagoMed Biopharma GmbH at the time of the study and LC holds shares in PhagoMed Biopharma GmbH and is the inventor on a patent application related to the work described here. The research of AS, AL, LVS and MV has been supported financially by PhagoMed Biopharma GmbH.
Funding This study was supported by the Austrian Research Promotion Agency (FFG).
Ethics approval The study was approved by the Charité ethical committee EA1 /088110.
Reformatted pdf to include figures in main text, for better readability.
References



