
ABSTRACT
In the current SARS-CoV-2 pandemic a key unsolved question is the quality and duration of acquired immunity in recovered individuals. This is crucial to solve, however SARS-CoV-2 has circulated for under five months, precluding a direct study. We therefore monitored 10 subjects over a time span of 35 years (1985-2020), providing a total of 2473 follow up person-months, and determined a) their antibody levels following infection by any of the four seasonal human coronaviruses, and b) the time period after which reinfections by the same virus can occur. An alarmingly short duration of protective immunity to coronaviruses was found by both analyses. We saw frequent reinfections at 12 months post-infection and substantial reduction in antibody levels as soon as 6 months post-infection.
ONE SENTENCE SUMMARY Coronavirus protective immunity is short-lasting
SARS-CoV-2 is a novel coronavirus responsible for an ongoing pandemic. Its rapid transmission is most probably caused by the fact that the virus entered a grossly naive, thus highly susceptible, human population, combined with the capacity of the virus to transmit during the asymptomatic phase of infection. Since no pharmaceutical interventions are universally available or applied, current policies to limit the spread of SARS-CoV-2 revolve around containment, social distancing, and the assumption that recovered patients develop protective immunity. It is still unclear whether protective immunity is indeed induced after infection, and for how long. The duration of protection will impact not only the overall course of the current pandemic, but also the post-pandemic period. To date, no concrete evidence of reinfection by SARS-CoV-2 is available, nor is any example of reinfection by SARS-CoV-1 or MERS-CoV, yet this is likely influenced by the recent emergence of SARS-CoV-2 and the limited scale of the SARS-CoV-1 and MERS-CoV epidemics. The limited available data on potential protective immunity against coronavirus reinfection is derived from one experimental infection study in volunteers (1), from which it is generally assumed that reinfection by coronaviruses can occur. Natural coronavirus infections and susceptibility to reinfection have not been investigated thus far. If they occur, reinfections will probably be dictated by two variables: exposure to the virus, and the quality of sustained immunity (2).
Even though it is not possible to investigate SARS-CoV-2 reinfections yet, the seasonal coronaviruses may serve as a reliable model. There are four species of seasonal coronaviruses, HCoV-NL63, HCoV-229E, HCoV-OC43, and HCoV-HKU1. All are associated with mostly mild respiratory tract infections. However, aside from being etiological agents of common cold, the four viruses are biologically dissimilar. Two belong to the genus Alphacoronavirus, and two to the genus Betacoronavirus. The viruses use characteristic receptor molecules to enter a target cell, and based on receptor distribution they do not all enter the same epithelial cell type in the lungs (3). Given this variability, the seasonal coronaviruses are the most representative virus group from which to conclude general coronavirus characteristics, particularly common denominators like host protective immunity and susceptibility to reinfection. Since most people experience their first seasonal coronavirus infection during early childhood (4-6), reinfections later in life can be investigated.
The aim of this study was to investigate the time period between coronavirus reinfections and the dynamics of antibody decline following infection. These parameters were assessed by measuring the immune response to each individual seasonal coronavirus, over an extended period. We followed healthy subjects for multiple decades at regular intervals. As IgG levels only increase after successful infection and—importantly—remain constant after unsuccessful viral challenge (1), increased antibody levels identify an infection. We used the carboxyl terminal of the nucleocapsid protein (NCt) as the target antigen in ELISA, as this protein is immunogenic, relatively low in interspecies conservation, yet high in intraspecies conservation when compared to the Spike protein (4, 7–10).
Frequency of infections and reinfections
From a prospective cohort study following adult males (see M&M and (11)), ten subjects were randomly selected. Follow-up of subjects started in 1985 and, besides a gap in follow up between 1997 and 2003, continued until 2020 at regular intervals (every 3 months prior to 1989, and every 6 months afterwards, Table 1). The cumulative period at which subjects were continuously followed totaled more than 200 person-years (2473 months). At start of the study, subject age ranged from 27 to 40 years; by the end of follow-up, subjects were 49 to 66 years old. The study was approved by the Medical Ethics Committee of the Amsterdam University Medical Center of the University of Amsterdam, the Netherlands (MEC 07/182).
Study subjects and seasonal coronavirus infections during follow up.
Infections by seasonal coronaviruses were defined as 1.4-fold increase in antibody levels occurring between two visits (1), and additionally the standard deviations of the mean at both visits not overlapping. A total of 132 events, ranging from 3 to 22 per subject, met these criteria and were thus considered infections (Table 1). The incidence rates of seasonal coronavirus reinfections per 100 person-years of follow-up was 15.3 (95% CI 10.0-23.4); 20.1 (95% CI 14.2-28.7); 16.4 (95% CI 10.426.1); 6.3 (95% CI 2.4-16.8), for HCoV-NL63, HCoV-229E, HCoV-OC43 and HCoV-HKU1 respectively. To test whether serological infection criteria represented symptomatic infections, we compared self-reported influenza like illnesses in the interval directly preceding the antibody peaks. Indeed, reporting of fever and cough were significantly associated (Fisher’s exact test p=0.031, supplementary table S1).
Median reinfection times of 33 (IQR 18 – 60), 31 (IQR 15 – 42), 27 (IQR 21 – 49), and 46 (IQR 36 – 68) months were found for HCoV-NL63, HCoV-229E, HCoV-OC43 and HCoV-HKU1, respectively, and 30 (IQR 18 – 54) months for all viruses combined (Fig. 1A). There was no statistically significant difference between the infection interval lengths of the individual viruses (Kruskal-Wallis test, P=0.74). In a few cases, re-infections occurred as early as 6 months (two times for HCoV-229E and one time for HCoV-OC43) and 9 months (two times for HCoV-NL63). The most frequent observed reinfection time was 12 months. For reinfections occurring as early as 6 months, we observed no reduction in antibodies between infections (Fig. 1A, white circles). At longer infection intervals, intermediate reductions in antibody levels were observed.
The ability to detect short-term reinfections is limited by the sampling interval. Importantly though, no reinfection was observed at the first subsequent follow-up visit after a 3 month interval (Fig. 1A). We did observe several reinfections at subsequent visits with a 6-month interval, suggesting that reinfections within 6 months do not occur. To further support this point, we examined ratios between antibody levels between two visits, both for 3 and 6 month follow-ups. As shown in Fig. 1B, only ratios below 1 were found in the observations with every 3 month-sampling, and we can therefore safely conclude that reinfections occur from 6 months on, yet most probably not earlier.

(A) The interval durations between reinfections. Only reinfections which were observed within a continuous follow-up period are shown. White dots indicate reinfections for which no intermediate decrease in antibody levels could be observed. Black vertical lines describe median reinfection times. (B) Changes in antibody levels post-infection relative to the follow-up interval duration. Each circle represents an infection. The x-axis describes the time until the next follow-up visit post-infection. The y-axis describes the change in antibody level at the subsequent visit. Larger circles represents a higher ratio-rise in antibody levels at the initial infection. The horizontal line indicates the border between increases (>1.0) or decreases (<1.0) in antibody levels at the next study visit (C) Kaplan-Meier curve showing decline of antibodies post infection (100%, 75% and 50%). The visit at which the infection was established was counted as time point 0, cases were subsequently followed. An event is defined when antibody levels drop below the indicated level. When the antibody level did not decrease to the indicated levels, an observation was censored at the last time point of active follow-up.
Antibody dynamics after infection
On some occasions, antibody rises reached high levels (up to ratio 11), represented in Fig. 1B as larger circles, yet these high values were never sustained at the next study visit (Fig. 1B). To investigate these dynamics further, we calculated the time until a 50%, 75%, or full return of antibody levels to baseline (pre-infection antibody levels). As can be seen in Fig. 1C, within 6 months post-infection the majority of people lost 50% of their antibodies, whereas after a year the majority lost 75%. A complete return to baseline levels occurred within 4 years for half of infections (Fig. 1C).
Simultaneous infections by alpha- and betacoronaviruses
Although our ELISA tests using the C-terminal part of the N protein were cautiously designed to be specific for each individual virus, we cannot rule out that a certain degree of antibody cross-reactivity occurred. We therefore investigated how often infections coincided, since cross-reactivity may have led to false labeling of infections. We observed no significant simultaneous infections for Alphacoronavirus (HCoV-NL63 or HCoV-229E) alongside Betacoronavirus (HCoV-HKUl or HCoV-OC43), however, we did see that infections by the betacoronaviruses HCoV-OC43 and HCoV-HKUl often coincided (38.5%, Table 2). Likewise, for alphacoronaviruses, HCoV-229E infections coincided with HCoV-NL63 infections in 59.5% of the cases, and vice versa in 44.9% of the cases. Hence, there is a risk that we overestimated the number infections and thus reinfections. We therefore re-analyzed the data with a more stringent definition of infection, including only the strongest antibody rise induced by a Betacoronavirus or Alphacoronavirus at a given time point. Under this definition we still found infection intervals comparable to the original data (supplementary Fig. S2), with minimum infection intervals as short as 6 months and frequent reinfections at 12 months, although the number of reinfections was obviously reduced.
Coinciding coronavirus infections
Broadly acting antibodies recognizing SARS-CoV-2
In theory, antibodies induced by (repeated) coronavirus infections may have broad coronavirus-recognizing characteristics. We therefore performed an additional ELISA on all 10 subjects, this time using the complete N protein of SARS-CoV-2, to allow detection of broadly reacting antibodies. Visual inspection suggested that broadly recognizing antibodies were produced, and were most likely induced by combined infections with HCoV-NL63 and HCoV-HKUl (subjects 2, 9 and 10, supplementary Fig. S1).
Coronavirus infections in changing seasons
To date it is uncertain whether SARS-CoV-2 will share the same winter prevalence peak that is observed for seasonal coronaviruses in non-equatorial countries. However, it is important to consider that winter preference of seasonal coronaviruses has only been determined by testing respiratory samples of people that experienced disease (12). Sampling and storage is therefore dictated by having symptoms and not by study protocol. If coronavirus spread continues unabated in summer, yet people rarely display symptoms (e.g. because of higher vitamin D levels) and are therefore not sampled, infections will remain undetected. Our serological study is unique because it avoids this sampling bias. The Netherlands has a typical temperate climate, and our study samples were collected at regular intervals. The sampling of each subject started at different times of the year, and, because of the 3 or 6 months regime of visits, samples were collected throughout all seasons. Consequently, we can for the first time visualize in an unbiased manner the seasonality of coronavirus infections. As shown in Fig. 2, May, June, July, August and September indeed show the lowest probability of infections for all four seasonal coronaviruses (Wilcoxon signed-rank test, p=0.005).
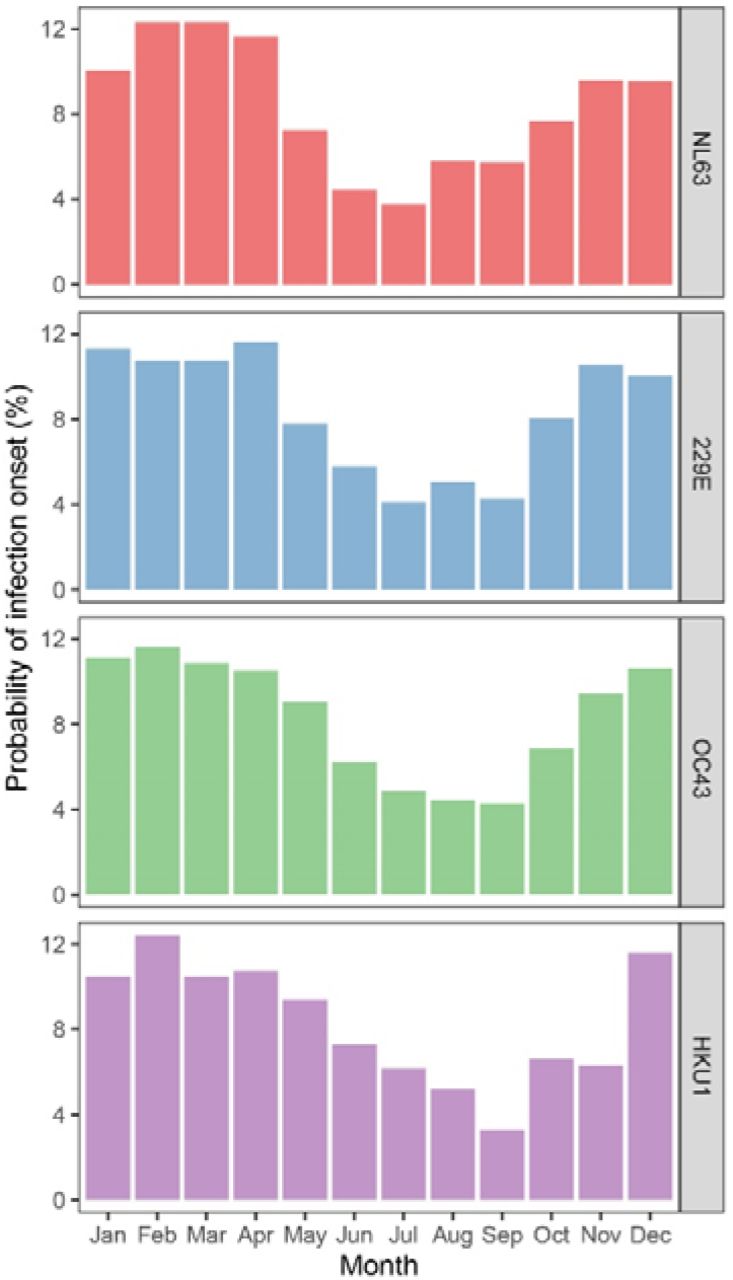
The prevalence of the four seasonal coronaviruses shown as the probability of infection onset for a specific month. The months between and including the months of the follow-up visits at which the infection was established and the prior follow-up visit were counted. The value per month was divided by the number of months between follow-up visits.
DISCUSSION
We show, for the first time, that reinfections with all seasonal coronaviruses occur in nature. The majority of reinfections occurred within 3 years. If SARS-CoV-2 will behave like a seasonal coronavirus in the future, a similar pattern may be expected. However, this time span between infections does not indicate that an individual’s protective immunity lasts for the same period of time, as reinfection is also dependent on re-exposure. In fact, based on the minimum infection intervals and the dynamics of antibody waning that we observed, the protective immunity may last as little as 6 to 12 months. Recently Kissler et al. modeled the protective immunity and reinfection dynamics HCoV-OC43 and HCoV-HKU1 and estimated a 45 week period of protective immunity (13). Our serological study confirms this prediction.
When we view our findings in light of the current control actions taken for SARS-CoV-2, it is clear that coronavirus reinfection risk is key to public health policy. Here we reveal a risk that in the near future, serology based tests that measure previous infections for SARS-CoV-2 may have limited use if that infection has occurred >1 year prior to sampling. Additionally, vaccine studies should anticipate that sustained protective immunity may be uncertain for coronaviruses, and repeated yearly or half-yearly vaccinations may be needed to circumvent ongoing transmission.
There is ongoing discussion regarding herd immunity for SARS-CoV-2 control. Herd immunity occurs when a threshold proportion of a population is immune to a certain pathogen, and protects even non-immune individuals against the infection by limiting overall spread. This effect has been observed for a variety of viruses, mainly those that are partially controlled via vaccination programs, such as hepatitis A virus (14), influenza A virus (15), and human papilloma virus (16). In the case of SARS-CoV-2, achieving herd immunity may be challenging due to rapid loss of protective immunity. It was recently suggested that recovered individuals should receive a so-called “immunity passport” (17) which would allow them to relax social distancing measures and provide governments with data on herd immunity levels in the population. However, as protective immunity may be lost by 6 months post infection, the prospect of reaching functional herd immunity by natural infection seems very unlikely.
It is generally assumed that exposure to each seasonal coronavirus is not equal. The Betacoronavirus HCoV-HKU1 is considered to have the lowest prevalence (4, 13) and we found the same. We also observed that antibody levels to HCoV-HKU1 were low over the whole range (all samples tested). Whether this indicates HCoV-HKU1 infection elicits a poor induction of antibodies is intriguing, yet not answerable in our study. We used a relatively small part of the N protein as the antigen in our tests, to most specifically identify infections by each virus individually. This small part of the protein may not contain the most immunodominant epitopes of HCoV-HKU1. A neutralization test would ideally be done to solve the issue of protective immunity raised by HCoV-HKU1; however, unfortunately no culturing system suitable for neutralization testing is available for HCoV-HKU1 (18).
We noticed three subjects to carry antibodies recognizing SARS-CoV-2 N protein at certain time points. It is unlikely that they had been infected with a SARS-CoV-2-like virus in 1985 (subject 10), 1992 (subject 2), or 2006 (subject 9), and we therefore suggest that broadly acting antibodies may have been induced by coinciding infections of an Alpha- and a Betacoronavirus (in our subject(s) HCoV-HKU1 and HCoV-NL63). To explore this finding we looked at the genetic distance and consequently amino acid differences in the structural protein of the various coronaviruses (supplementary Table S2). Notably, SARS-CoV-2 N protein has only 32% and 34% identity on the amino acid level with the N protein of HCoV-OC43 and HCoV-HKU1 respectively, and only 26% and 24% identity with HCoV-NL63 and HCoV-229E respectively. Similarly, the distance between Alphacoronavirus and Betacoronavirus N protein is large (only 24% to 26% amino acid identity). Still, we cannot exclude the presence of conserved (conformational) epitopes in HCoV-HKU1 and HCoV-NL63 N protein that may result in a more broadly acting antibody response, due to simultaneous exposure in concurrent infections. Additional screening, including more subjects, is required for confirmation.
Our study was subject to limitations. One is the inability to sequence the virus genome during infection. In theory, strain variation could play a role in susceptibility to reinfection. HCoV-NL63, HCoV-OC43, and HCoV-HKU1 all show different co-circulating genetic clusters (19-21). For HCoV-229E, there are no major genetic subtypes known, like the current SARS-CoV-2 situation (22, 23). It could be that our observation of short-term immunity is influenced by genotype variation, and a study on protective immunity would therefore ideally allow sequencing of re-infecting strains from respiratory material; however, this is intractable in a natural infection study because virus shedding in reinfections can be as short as one day (1). Another limitation is that the studies subjects were all males. For COVID-19, and also HCoV-NL63, men have a higher incidence of disease (25,26), and it is therefore of interest to determine the dynamics of protective immunity also in a cohort of healthy women.
In conclusion, seasonal human coronaviruses have little in common, apart from causing common cold. Still, they all seem to induce a short-lasting immunity with rapid loss of antibodies. This may well be a general denominator for human coronaviruses.
Data Availability
All data are available upon request from the authors
Author contributions
A.W.D.E: Conceptualization, Writing – original draft, review and editing, Investigation, Visualization, Formal Analysis, Validation;J.K.: Investigation, Writing – original draft, review and editing, A.C.R.H.: Resources; M.B.: Investigation, Resources; M.K.: Investigation; M.F.J.: Investigation, Methodology; A.M.: Formal Analysis; C.M.K.: Writing – original draft, review and editing, Formal Analysis; P.R.: Resources; M.P.: Resources; P.S.: Resources; M. D.: Investigation, Methodology; L.v.d.H.: Conceptualization, Writing – original draft, Writing – review and editing, Supervision.
Competing interests
Authors declare no competing interests.
Acknowledgments
The authors gratefully acknowledge the Amsterdam Cohort Studies (ACS) on HIV infection and AIDS, a collaboration between the Public Health Service of Amsterdam, the Amsterdam UMC of the University of Amsterdam, Sanquin Blood Supply Foundation, Medical Center Jan van Goyen, and the HIV Focus Center of the DC-Clinics. It is part of the Netherlands HIV Monitoring Foundation and financially supported by the Center for Infectious Disease Control of the Netherlands National Institute for Public Health and the Environment. The authors thank all ACS participants for their contribution, as well as the ACS study nurses, data-managers, and lab technicians. This work was supported by a grant from the European Union’s Horizon 2020 research and innovation programme, under the Marie Sktodowska-Curie Actions grant agreement no. 721367 (HONOURs), Amsterdam UMC funding connected to HONOURs, and the Amsterdam UMC PhD scholarship of A.W.D. Edridge.




{kind=link}
{kind=link}