
ABSTRACT
Background The extent of spread of SARS coronavirus 2 (SARS-CoV-2) in the UK and elsewhere is unknown because typically only symptomatic individuals are diagnosed. We performed a serological study of recent blood donors in Scotland to detect antibodies to SARS-CoV-2 as a marker of past infection.
Methods A pseudotyped SARS-CoV-2 virus microneutralisation assay was used to detect neutralising antibodies to SARS-CoV-2. The study group comprised samples from 1000 blood donors collected in Scotland during March, 2020. Controls were collected from 100 donors in Scotland during 2019.
Findings All samples collected on the 17th March, 2020 (n=500) were negative in the pseudotyped SARS-CoV-2 virus microneutralisation assay. Neutralising antibodies were detected in 5 of the 500 samples collected 21st – 23rd March; one further sample was reactive in an anti-spike ELISA.
Interpretation Although we cannot use the rise in numbers seropositive to infer the contemporary seroprevalence or the growth rate of the epidemic, we note that they are consistent with frequency of reported diagnosed infections and SARS-CoV-2-associated deaths reported in that time period in Scotland, given that seroconversion takes up to 2-3 weeks. It should also be noted that blood donors are not representative of the general population; in particular, those with a history of recent respiratory infections are deferred. Finally, it is unknown what proportion of infected individuals seroconvert and become reactive in the assays used. Serial follow up studies are needed to track infection and seroconversion in this and other similar populations However, these data indicate that sero-surveys of blood banks can serve as a useful tool for tracking the emergence and progression of an epidemic like the current SARS-CoV-2 outbreak.
INTRODUCTION
SARS-coronavirus-2 (SARS-CoV-2) emerged in late 2019 in Hubei province China as a cause of respiratory disease occasionally leading to acute respiratory distress syndrome and death (COVID-19)1-4. Since the first reports in December, 2019, infections with SARS-CoV-2 have been reported from an increasing number of countries worldwide, with particularly high incidence of diagnosed infections and associated deaths from respiratory disease initially in China but more recently in Italy, Iran, Spain, France and the USA (https://www.who.int/emergencies/diseases/novel-coronavirus-2019). Increasing age, male gender, smoking and comorbidities such as cardiac disease, hypertension and diabetes have been identified as risk factors for severe infections5-7. For as yet unknown reasons, infants and children seem to be less at risk for moderate to severe COVID-19 disease8.
Compared to Italy and Spain, the SARS-CoV-2 pandemic was at a relatively early stage in the UK in early March 2020. However, a dramatic rise in the number of admissions of patients with severe SARS-CoV-2 infections followed and emergency plans have been implemented. Prediction of the future severity of the outbreak, and most specifically the trajectory of severe cases that require hospitalisation and intensive care support is key to the national response, as it is throughout several affected countries worldwide. Predicting disease outcomes is complex; in addition to the basic information of the proportions of individuals in different age ranges who develop severe disease, the severity of the outbreak is also crucially dependent on current population immunity and virus transmissibility. Mean estimates of R0 of 3.29 (2.12 – 4.45 interquartile range from 12 studies based on presumed SARS-CoV-2 naive populations)9 indicate that a 50%-75% minimum level of herd immunity from past infection is sufficient for a sustained reduction of new infections with time. SARS-CoV-2 spread may be further influenced by seasonal changes in transmissibility, as observed for other respiratory coronaviruses infecting humans10-12.
In the current study we have taken the first steps towards estimating SARS-CoV-2 exposure in a European country by measuring seroprevalence in a sample of blood donations. Samples from donors in an age range from 18-75 years collected across Scotland on the 17th March and 21st-23rd of March, 2020 were assayed for neutralising antibody to SARS-CoV-2 using a pseudotyped SARS-CoV-2 virus microneutralisation (pMN) assay format used previously for SARS-CoV-1 and Ebolavirus seroepidemiology purposes13-15 and confirmed using an enzyme-immunosorbant (ELISA) assay detecting antibodies to spike protein. The detection frequency of neutralising antibodies and a discussion of its applicability for estimating population level exposure are presented.
METHODS
Samples
A total of 500 plasma samples collected on the 17th March and a further 500 on the 21st-23rd March, 2020 were analysed in the study. To serve as negative controls, 100 blood donor samples were tested in parallel from the Scottish National Blood Transfusion Service (SNBTS) anonymous archive collected between September 2018 and December 2019 (IRAS Project No. 18005), before the first reports of the spread of SARS-CoV-2 in China. Donor selection criteria are described in Suppl. Data. SNBTS blood donors give fully informed consent to virological testing, donation was made under the SNBTS Blood Establishment Authorisation and the study was approved by the SNBTS Research and Sample Governance Committee. Seven control samples from contract-traced individuals who were PCR-confirmed as SARS-Cov-2 infected were used as positive controls in the study. All the individuals from whom the positive control sera samples were taken had asymptomatic SARS-CoV-2 infections and recruited through the ISARIC WHO Clinical Characterisation Protocol UK (CCP-UK) at the discharge plus 28 day time-point. Samples were heat inactivated prior to serological testing by incubation at 56°C for 30 minutes.
SARS-CoV-2 pseudotype microneutralisation (pMN) assay
A lentivirus-based SARS-CoV-2 pseudovirus particle was constructed displaying the full spike protein on the surface of the pseudotyped virus using a synthetic codon optimised SARS-CoV-2 expression construct (Accession number: YP_009724390.1). The methodology has previously been used to produce pseudotyped viruses for SARS-CoV-1, MERS-CoV and Ebolavirus13-15. Virus infectivity was determined by titration on HEK 293T ACE2-plasmid transfected cells as previously described (Nie et al, 2020). Neutralizing antibody (Nab) titres were determined by endpoint two-fold serial dilutions of test samples mixed with 105 relative light units (RLU) of pseudotyped virus, incubated at 37°C for two hours and then mixed with 104 HEK 293T ACE2-transfected cells per well. Plates were incubated for 72 hours at 37°C and then cells were lysed and assayed for luciferase expression. Neutralization titres are expressed as IC50values.
During the assay, plates were barcoded and controls were spaced throughout the runs. Individuals were blinded regarding the arrangement of spaced positive controls on the plates.
Enzyme-linked immunosorbent assay (ELISA)
Antibodies to the trimeric S protein were detected by ELISA. MAXISORP immunoplates (442404; NUNC) were coated with StrepMAB-Classic (2-1507-001;iba). Plates were blocked with 2% skimmed milk in PBS for one hour and then incubated with 0.125ug of soluble trimeric SARS-CoV2 trimeric Spike protein or 2% skimmed milk in PBS. After one hour, plasma was added at 1:50 dilution, followed by ALP-conjugated anti-human IgG (A9544; Sigma) at 1:10,000 dilution or ALP-conjugated anti-human IgM (A9794; Sigma) at 1:5,000 dilution. The reaction was developed by the addition of PNPP substrate and stopped with NaOH. The absorbance was measured at 405nm after 1 hour.
Statistical and Modelling Analysis
Samples were analysed following the protocol outlined in Ferrara & Temperton16 which was implemented using the SciPy curve fit15. Values for each plate (V) were corrected for background signal by subtracting the average of 6 negative control wells 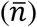 . The standardised percentage neutralisation for each sample value (S) was calculated by benchmarking against the average of 6 technical positive controls for each plate
. The standardised percentage neutralisation for each sample value (S) was calculated by benchmarking against the average of 6 technical positive controls for each plate 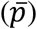 using the equation:
using the equation:
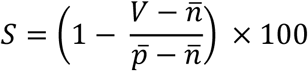 Standardised percentages from two replicates of the same sample were averaged for each dilution. A three-parameter logistic function was then fit to these averages using non-linear least squares:
Standardised percentages from two replicates of the same sample were averaged for each dilution. A three-parameter logistic function was then fit to these averages using non-linear least squares:
 Here, the averaged standardised neutralisation percentage
Here, the averaged standardised neutralisation percentage 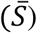 , is a function of the logarithm (base 10) of the dilution factor (D)and the 50% inhibitory concentration (IC50). The IC50value corresponds to the dilution factor where 50% neutralisation is predicted to be achieved. Larger IC50values correspond to samples with stronger antibody responses which require higher dilutions to reduce neutralisation. The parameter a describes the asymptote of the curve and was fixed to 100 (100% neutralisation was the maximum). The parameter C describes the steepness of the slope for the sigmoidal curve and was restricted to take values < 0 (only negative responses of neutralisation to dose were permitted). The quality of the model fit was assessed using the summary statistic, R2, which examines the differences between the fitted values of the logistic function and the data points. A larger value of R2 corresponds to a higher proportion of the variance explained (compared to a model describing all of the neutralisation values using a single mean). Some positive controls were repeated across different plates, so for these an error-weighted mean of IC50was calculated from values estimated from separate replicates.
, is a function of the logarithm (base 10) of the dilution factor (D)and the 50% inhibitory concentration (IC50). The IC50value corresponds to the dilution factor where 50% neutralisation is predicted to be achieved. Larger IC50values correspond to samples with stronger antibody responses which require higher dilutions to reduce neutralisation. The parameter a describes the asymptote of the curve and was fixed to 100 (100% neutralisation was the maximum). The parameter C describes the steepness of the slope for the sigmoidal curve and was restricted to take values < 0 (only negative responses of neutralisation to dose were permitted). The quality of the model fit was assessed using the summary statistic, R2, which examines the differences between the fitted values of the logistic function and the data points. A larger value of R2 corresponds to a higher proportion of the variance explained (compared to a model describing all of the neutralisation values using a single mean). Some positive controls were repeated across different plates, so for these an error-weighted mean of IC50was calculated from values estimated from separate replicates.
RESULTS
Serology testing
Sera from 1000 blood donors and 100 pre-pandemic controls (from 2019) were assayed for anti-SARS-CoV-2 neutralising antibody by pMN assay. Test samples comprised plasma collected from randomly selected donors on the 17th March (n=500), the 21st-23rd March (n=500), 2020 (Figure 1) and prior to the epidemic obtained between September, 2018 and December 2019 (n=100). Blood donor samples were collected across Scotland with representation of both primarily urban (eg. Edinburgh Health Board) and rural (Inverness Health Board) areas. There was an approximately equal representation of males and females amongst donors but a restricted age range from 18 (minimum donor age) to 75 (Fig. 1); this was also skewed towards older age ranges compared to the Scottish population within that range (Fig. S1; Suppl. Data).
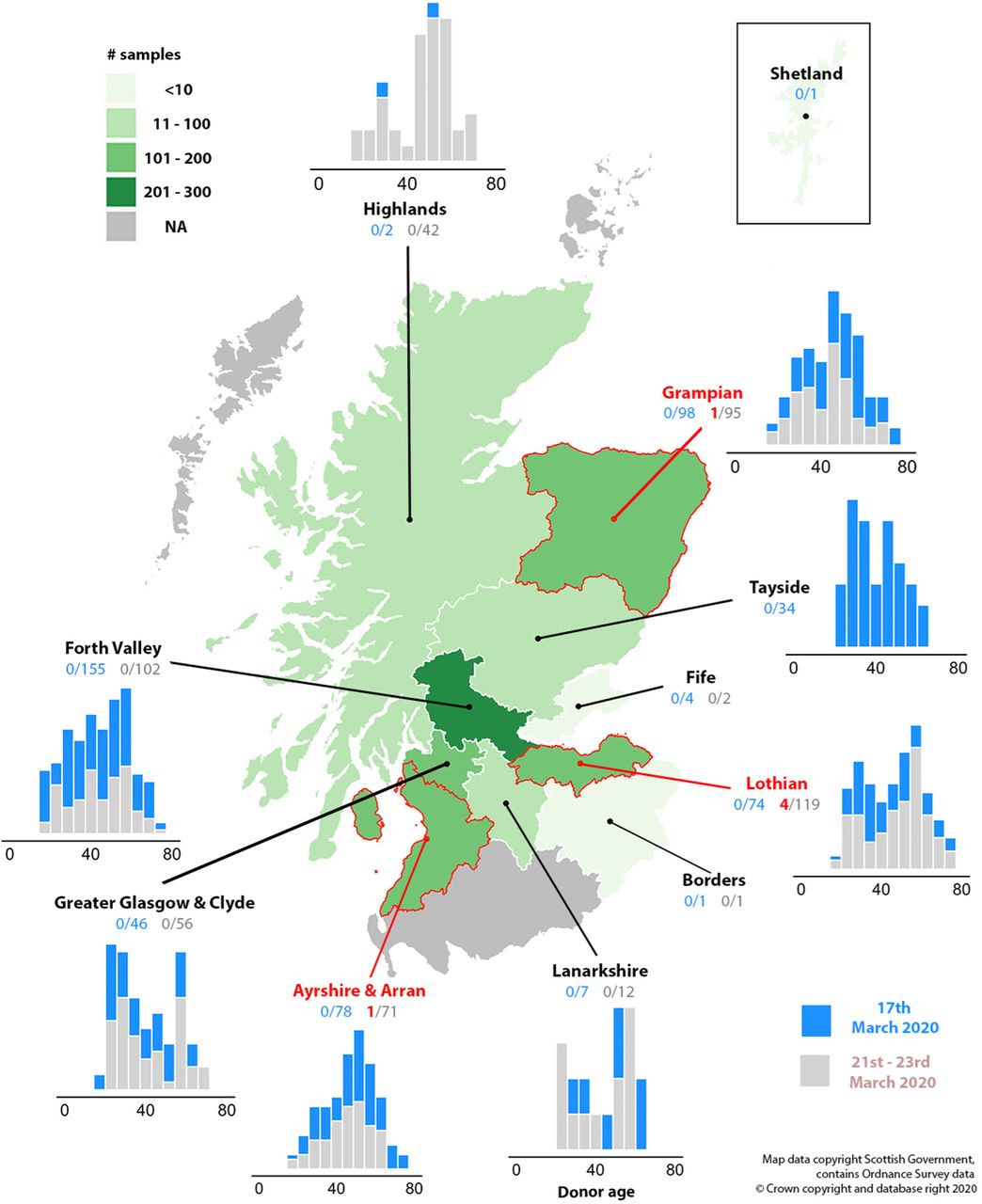
SAMPLING DISTRIBUTION OF THE SCOTTISH BLOOD DONORS
The numbers of seropositive blood donor samples in the study divided regionally by health board. Sampling density is indicated by the shading of each region. The locations of the seropositive donors are indicated in red text, arrows and borders. Histograms show the sampling frequencies of different age classes for week 1 (blue) and week 2 (grey) for each health board.
Samples from both weeks in 2020 and the 2019 controls were assayed by pMN assays using a two-fold dilution series of plasma from 1:20 – 1:640. The reduction in luciferase signal through antibody binding at each dilution was curve-fitted in order to estimate the dilution leading to a 50% inhibition of pseudotype replication (IC50). A second metric is the degree of fit of the datapoints from the dilution series to the predicted sigmoid curve using for fitting, expressed as R2. Samples showing non-specific blocking of entry (ie. non-concentration dependent) will show a low R2 value. Of the 7 positive control samples, 6 possessed detectable neutralising ability, above the range seen in the pre-pandemic samples, and good curve fits (IC50 range 83.18-323.59 R2 range: 0.79-0.96; Fig. 2). A total of 5 blood donor samples from the second week showed evidence for neutralising antibody (IC50 range 78.42-3434.90; R2 range: 0.64-0.95). All other samples from 2020 showed low IC50 values, low R2 values or in most cases low values for both and were classified as negative.
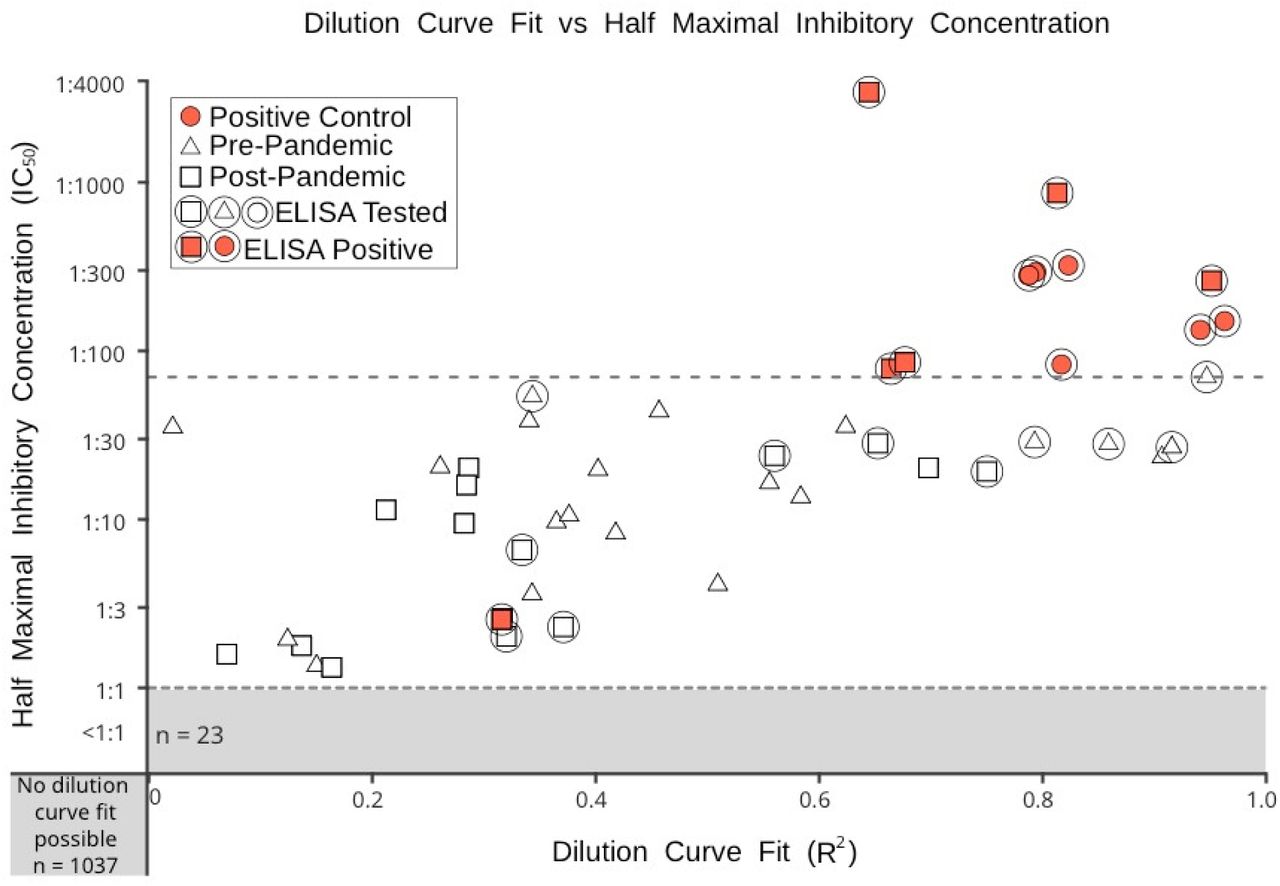
ASSOCIATION OF IC50 (NEUTRALISING ANTIBODY) VALUES AND R2 (GOODNESSS OF CURVE FITS) OF THE TEST SAMPLES
A plot of R2 and IC50values for blood donor and positive control samples. Triangular icons denote pre-pandemic samples, square icons indicate post-pandemic samples, red circles indicate the positive control samples. Anti-spike protein ELISA tested samples are circled. Post-pandemic positive ELISA tested samples are indicated by by filled red squares.. The dotted line indicates the highest IC50 value found among the pre-pandemic samples.
To verify the antibody results, the positive samples and a selection of negatives, including those close to the threshold of positivity in the pMN assay, were tested in an anti-spike protein ELISA for IgG and IgM antibodies at a single 1:50 dilution. ELISA analysis identified a further antibody positive test sample that was not clearly positive in the pMN assay (IC50: 2.55 R2:0.32; lower left quadrant in Fig. 2). Optical densities (ODs) recorded for the positive controls and blood donor samples that were positive in the pMN assay showed a good correlation with neutralising antibody titres in the ELISA for IgG antibodies (Fig. 3). The two weak positive samples in the pMN assay were also clearly reactive in the ELISA and have been assigned as positive in this study. All other samples were negative. Low or undetectable levels of IgM were detected by ELISA (data not shown).

{kind=link}
{kind=link}
{kind=link}
RELATIONSHIP BETWEEN IC50 AND ELISA REACTIVITY OF POSITIVE SAMPLES
A plot of neutralising antibody IC50value vs OD of ELISA samples in an IgG anti-spike protein ELISA. Post-pandemic ELISA positive samples are denoted by a red square with a red circle surrounding it. Positive controls are denoted with a filled red circle.
Overall, 6 donor samples from the March 21st – 23rd time point were positive for anti-SARS-CoV-2. All samples from donors in the previous week and in 2019 were negative. Positive donors originated from the Edinburgh Health Board area (n=4), Grampian (n=1) and Ayrshire & Arran (n=1) with an age range from 20-60 years.
DISCUSSION
This study reports the prevalence of neutralising antibodies in blood donations given over two weeks in mid-March 2020 in two regions of Scotland, a period when clinical cases were beginning to present; an estimated 25 deaths from SARS-CoV-2 infections had been recorded in Scotland at the time of sample collection. In interpreting the significance of this seroprevalence, we review several factors which may influence antibody detection rates and how this might translate into calculations of population exposure and immunity and projections of the outcomes of the SARS-CoV-2 outbreak.
The representativeness of the study region
The study was based upon testing anonymised samples from blood donors collected in March, 2020. Scotland has a population of 5.4 million and a relatively low population density (67.2/km2). However, it contains several large cities (Glasgow, Edinburgh) which geospatially would support transmission networks typical of much of Western Europe. Public health data for COVID-19 cases indicated that by the 26th March, shortly after donor collection, a total of 894 infections had been diagnosed and 25 deaths recorded (normalised incidence: 0.46/100,000). This compares to an overall confirmed incidence for the UK of 0.62/100,000 on the same day. The rate in Scotland was higher than Germany (0.18/100,000), comparable to Denmark (0.41/100,000), France (0.30/100,000) and Spain (0.42/100,000) but substantially lower than Italy (11.1/100,000). The value of these comparisons has to be tempered by potential differences between countries in criteria used to attribute death from SARS-CoV-2 infection. Furthermore the situation is highly dynamic – on the 1st April, the rate in Scotland had risen to 0.87/100,000 but it was much higher in England (2.5/100,000) and rates may diverge further in the future.
The representativeness of blood donors
The demographics of blood donors differs in several aspects from the general population, most notably because of the exclusion of those at risk for blood-borne viruses (HIV-1, HCV, HBV) and syphilis. However, they might be considered a reasonable representation of the adult population for SARS-CoV-2 (and other respiratory viruses) in the absence of any obvious predisposing factors for infection. The only other general exclusion was a 4 week donation deferral period in those recently travelled to specified countries at risk for arbovirus and malaria infections (including China). However, since the onset of the outbreak, donors have also been deferred if they have a history of a laboratory confirmed SARS-CoV-2 diagnosis, or respiratory symptoms consistent with SARS-CoV-2 infection within the previous 28 or 14 days (listed in Suppl. Data). The potential exclusion of infected symptomatic donors will therefore systematically reduce the seroprevalence recorded and skew the estimate of population seroconversion. The extent to which this occurs depends on the outcomes of infection. Amongst many data sources, a comprehensive analysis published by China CDC17 estimates that 81% of those infected will have mild or inapparent disease (symptoms of upper respiratory tract viral infection, mild fever, cough (dry), sore throat, nasal congestion, malaise, headache, muscle pain but without dyspnoea or other sign of respiratory distress or insufficiency). Given that these symptoms do not differ substantially from other winter respiratory infections, it is likely that a substantial proportion of the estimated 81% of mild cases would proceed to donate.
Sensitivity and specificity of the serological tests used
There are currently no agreed standards or controls available for serological testing for SARS-CoV-2. The results presented in the study are therefore based on a formally non-validated assay. However, we believe that the neutralisation antibody test is likely to be robust. Firstly, it is based on the same design as pMN assays for SARS-CoV-1, ebolavirus and influenza A virus; pMN assays that have demonstrated high specificity for target virus neutralising antibodies and a sensitivity that is often greater than achieved in neutralising antibody assays for whole virus13-15. Overall, although sampling was limited, there was concordance between the pMN assay to detect neutralising antibody and the detection of anti-spike IgG antibody by ELISA. The pMN assay does, however, require comparison with neutralisation of live virus for further interpretation of these data.
The specificity of both assays is potentially influenced by cross-reactivity with other human coronaviruses, including the respiratory viruses, OC43 and HKUI in the Betacoronavirus genus that circulate in winter months. Some previous studies have indicated an absence of reactivity of negative control sera against the SARS-CoV-1 spike protein in ELISA and the high specificity of assays based on the spike protein compared to those using the more conserved nucleoprotein or whole virus18. SARS-CoV-2 spike protein is likely to share this specificity. Existing data on neutralising antibodies indicates that assay specificity may be even greater; although both SARS-CoV-1 and −2 enter cells through the ACE-2 receptor19-21, and possess structurally similar spike proteins20,22-24, it has recently been demonstrated that neutralising monoclonal antibodies (MAbs) against SARS-CoV-1 infection bound but did not neutralise SARS-CoV-225, Conversely, MAbs raised against SAR-CoV-2 showed little or no cross-neutralisation of SARS-CoV-126,27. However, cross-reactivity is most unlikely to cause an assay specificity problem, as SARS-CoV-1 has never spread significantly in the UK or elsewhere in Europe28.
The relationship between seroprevalence and SARS-CoV-2 exposure and infection
Serological tests used for determining population exposure are based upon an assumption of a durable virus-specific IgG antibody response to infection that persists for years after infection. This is typically assumed in seroprevalence studies for many human pathogens, including poliovirus, measles, and hepatitis B virus. There is little information, however, on the frequency, time course and durability of the antibody response to SARS-CoV-2 on infection and the extent to which this might vary in those who suffered different disease severities; these are all factors potentially influencing the effectiveness of serological tests to detect past infections 11. An investigation of immune responses in patients in the related MERS-CoV indicate that antibody seroconversion in coronavirus infections may not occur in all individuals29. Of concern, those with mild MERS-CoV infections (asymptomatic, or symptoms restricted to fever, headache, cough, and malaise) invariably failed to seroconvert for IgM or IgG antibody in immunoassays and similarly failed to mount a neutralising antibody throughout the study period. These observations contrast with universal seroconversion in those with moderate or severe MERS. If SARS-CoV-2 showed a similar disease-related differences in serological responses, it is possible that a substantial proportion of the test population in the current study may have been infected but negative for anti-SARS-CoV-2 neutralising antibodies.
Studies on SARS-CoV-1 showed that most patients seroconverted after infection but this was delayed (17-21 days) compared to a typical time course for seroconversion with other respiratory viruses30,31. Several studies demonstrate seroconversion of IgG antibodies against the spike protein 1-3 weeks after infection with SARS-CoV-2 32-36. A robust early and strong IgM, IgG and neutralising antibody responses was similarly observed in hospitalized individuals identified on contact tracing - in 50% of subjects by day 7, and in all nine by day 14 (https://doi.org/10.1038/s41586-020-2196-x). A proteome analysis of antibody induced by SARS-CoV-2 in 29 donors revealed a reproducibly detectable response to Spike and N proteins following SARS-CoV-2 infection37. However, there remains considerable uncertainty in this area. For example, a study of 175 hospitalized patients in China indicated a wide range of neutralizing antibody levels detectable post-recovery, with a some below the level of detection, especially amongst younger patients (9/55 assayed in those <40 years old)38. Similarly, the median time for seroconversion to IgG in hospitalized patients was found to be 14 days in a study of 173 patients in China, with some taking up to 1 month to generate antibody detectable in a commercial ELISA test39. Technical considerations aside, the temporal gap between exposure and seroconversion, coupled with the rapid spread of this virus means that during the initial phase of the epidemic an important difference will exist - at any given time point - between the fraction of those recently exposed/infected and the fraction of those who have seroconverted
Conclusions
The raw seroprevalence estimates obtained in the current study of blood donors in Scotland were 0/500 and 6/500 at two sampling points in mid-March 2020. These estimates reflect the prior exposure to SARS-CoV-2 in this blood donor population over the preceding weeks. Extrapolating this figure to represent the past infection frequency of SARS-CoV-2 in this and the wider community has a number of uncertainties that need to resolved in future studies in this and other similar populations. However, this study implements a novel method of tracking the spread of epidemics like the current SARS-CoV-2 outbreak, and should be carefully considered as a routine component of an immediate response to such an impending threat.
AUTHORS’ CONTRIBUTIONS
Thompson, C., Chinnakannan, S., Dejnirattisai, W., Edmans, M., Fyfe, A., Kooblall, K., Lee, L., Lim, N., Liu, C., López-Camacho, C., Mongkolsapaya, J., Odon, V., Ramamurthy, N., Ratcliff, J., Supasa, P., Wang, B. and Mentzer, A., performed the sample acquisition, laboratory testing and reporting of the pseudotype and ELISA testing. Imlach C., McInally C., Harvala, H. and Jarvis, L.M. established the sample sets, archiving and data provision of the samples used in the study. Grayson, N., Lourenco, J., Penman, B.S., Semple, M.G., Baillie JK, and Paton, R.S. performed the data analysis and results interpretation. Turner, M., Thompson, C., Temperton, N., Gupta, S., Klenerman, P., Screaton, G.R. and Simmonds, P conceived and designed the specifics of the study, the data interpretation and drafting of the manuscript. All co-authors contributed to the editing and final drafting of the manuscript and figures.
CONFLICT OF INTEREST STATEMENTS
The authors declare no conflict of interest
ROLE OF FUNDING SOURCE
The funders played no role in the design, execution or reporting of the study.
ETHICS COMMITTEE APPROVAL
Ethical approval for the SNBTS anonymous archive - IRAS Project No. 18005. SNBTS blood donors give fully informed consent to virological testing, donation was made under the SNBTS Blood Establishment Authorisation and the study was approved by the SNBTS Research and Sample Governance Committee.
FUNDING
This work was supported by the Medical Research Council [grant number MC_PC_19059]. National Institute for Health Research Biomedical Research Centre Funding Scheme (to G.R.S.), the Chinese Academy of Medical Sciences (CAMS) Innovation Fund for Medical Science (CIFMS), China (grant number: 2018-I2M-2-002). PK, PS and G.R.S. is supported as a Wellcome Trust Senior Investigator (grant 095541/A/11/Z; WT109965/MA). PK is an NIHR Senior Investigator. NG was supported via grant to Philip Goulder (WTIA Grant WT104748MA) and a grant to John Frater (Medical Research Council MR/L006588/1). CPT was funded by an ERC research grant ‘UNIFLUVAC’ and two MRC CiC grants (Ref: BR00140).
ACKNOWLEDGEMENTS
We would like to acknowledge the help and collaboration of many SNBTS staff for provision and preparation of samples (anonymous archive and recent donation samples). We acknowledge the wider support of ISARIC4C.
REFERENCES




Subject Area
Reviews and Context
0
Comment
0
TRIP Peer Reviews
0
Community Reviews
0
Automated Services
19
Blogs/Media
Author Videos