
ABSTRACT
3,4-methylenedioxymethamphetamine-assisted therapy (MDMA-AT) for post-traumatic stress disorder (PTSD) has demonstrated promise in multiple clinical trials. MDMA is hypothesized to facilitate the therapeutic process, in part, by decreasing fear response during fear memory processing while increasing extinction learning retention. The acute administration of MDMA in healthy controls modifies recruitment of brain regions involved in the hyperactive fear response in PTSD such as the amygdala and hippocampus. However, to date there have been no neuroimaging studies aimed at directly elucidating the neural impact of MDMA-AT in PTSD patients. We analyzed brain activity and connectivity via functional MRI during both rest and autobiographical memory (trauma and neutral) response before and two-months after MDMA-AT for PTSD in nine veterans and first-responders. We find that MDMA-AT (i) increases amygdala-hippocampal resting-state functional connectivity, and (ii) reduces amygdala-precuneus functional connectivity during neutral autobiographical memory recall in a manner that co-varies with reduction of PTSD severity. These findings compliment previous research indicating that amygdala-hippocampal functional connectivity is a potential target of MDMA-AT and highlights other regions of interest related to memory processes. More research is necessary to determine if these findings are specific to MDMA-AT compared to other types of treatment for PTSD.
1 INTRODUCTION
Post-traumatic stress disorder (PTSD), which can arise following exposure to a traumatic event or repeated stressful events, impacts hundreds of millions of people, and is a debilitating social and economic burden on individuals and their families. PTSD is associated with an increased fear response (VanElzakker, Staples-Bradley, and Shin 2018) and distressing and intrusive re-experiencing of traumatic memories (Ehlers 2010) that often serves as a barrier to the therapeutic process. Current psychological therapies for PTSD have high dropout rates (Goetter et al. 2015), especially trauma focused therapies (Lewis et al. 2020). When presented with trauma-related stimuli, PTSD patients have shown increased activation in the precuneus, retrosplenial cortex, anterior cingulate cortex (ACC), and amygdala compared to neutral stimuli (Sartory et al. 2013), suggesting augmented recruitment of brain regions involved in self-referential processing (Cavanna and Trimble 2006), salient autobiographical memory (Svoboda, McKinnon, and Levine 2006; Spreng, Mar, and Kim 2009; Sestieri et al. 2011; Maddock 1999), and fear and emotion (LeDoux 2003).
3,4-methylenedioxymethamphetamine-assisted therapy (MDMA-AT) is hypothesized to reduce the fear response associated with re-experiencing traumatic memories, and therefore may facilitate tolerable processing of traumatic content in patients with PTSD (Mithoefer et al. 2011). Phase 2 and 3 trials have demonstrated promise for MDMA-AT as a viable treatment for PTSD (Mitchell et al. 2021; Mithoefer et al. 2019; Wang et al. 2021; Jerome et al. 2020). In healthy individuals, acute administration of MDMA has been shown to enhance positive and reduce negative affect during the recollection of autobiographical memories, while preserving vividness and emotional intensity (Carhart-Harris et al. 2014). In another study, MDMA was found to preserve the memory accuracy when administered during both encoding and retrieval phases, while attenuating the recollection of salient details for both positive and negative memories, suggesting that MDMA alters emotional memory representations (Doss et al. 2018). Again in healthy controls, MDMA was found to enhance fear extinction retention rates compared to placebo when administered during extinction training phases (Maples-Keller et al. 2022). These findings suggest that MDMA may aid the therapeutic process, in part, by enabling patient access to emotionally challenging material and facilitating memory reconsolidation/fear extinction processes (Feduccia and Mithoefer 2018).
The exact neurobiological underpinnings of PTSD and the specific effects of MDMA-AT in individuals with PTSD have not been fully characterized, but several studies suggest the amygdala and hippocampus play an important role. The amygdala is broadly associated with fear response, and the hippocampus, associated with learning and memory, may provide contextual information necessary for cognitive-affect during memory recall (LeDoux 2003; Harnett, Goodman, and Knight 2020; Pitman et al. 2012). Sripada et al. (2012) found combat veterans with PTSD have decreased amygdala-hippocampal resting-state functional connectivity (RSFC) compared to combat veterans without PTSD, which the authors speculate may represent an inability to contextualize affective information in PTSD. In healthy volunteers, the acute administration of MDMA has the reverse effect, namely, increased amygdala-hippocampal RSFC (Carhart-Harris, 2015). Despite evidence that RSFC between the amygdala and hippocampus is implicated in PTSD and that this connection may be modulated by MDMA, no study to date has shown relationships between changes in these regions’ functional connectivity and the therapeutic effects of MDMA-AT.
Herein, we describe results from a study of combat veterans and first-responders undergoing MDMA-AT for PTSD in a randomized, double-blind, dose-response phase 2 clinical trial (Mithoefer et al. 2018). Both resting-state and task-fMRI data, acquired while individuals listened to a trauma-related or neutral script, were collected before and two months after MDMA-AT (follow-up scans were collected after the blind was broken). Prior to analysis, we hypothesized that MDMA-AT would increase RSFC between the amygdala and hippocampus (Sripada et al. 2012; Carhart-Harris et al. 2015). We further hypothesized that, at baseline, brain activity would be higher during the trauma-related listening task compared with the neutral listening task in regions associated with autobiographical memory, fear, and emotion, such as the precuneus, retrosplenial cortex, ACC, and amygdala, and that this effect would be reduced post-treatment (Sartory et al. 2013). Finally, we hypothesized that the magnitude of the pre-to-post treatment change in the functional connectivity of the amygdala and hippocampus would be correlated with the magnitude of change in the pre-to-post treatment Clinician-Administered PTSD Scale (CAPS-IV) total severity scores (Blake et al. 1990). Understanding the neurobiological mechanisms of PTSD and how we may modulate these mechanisms is imperative to developing effective, personalized treatments to reduce the burden of this disease.
2. METHODS
2.1 Trial design
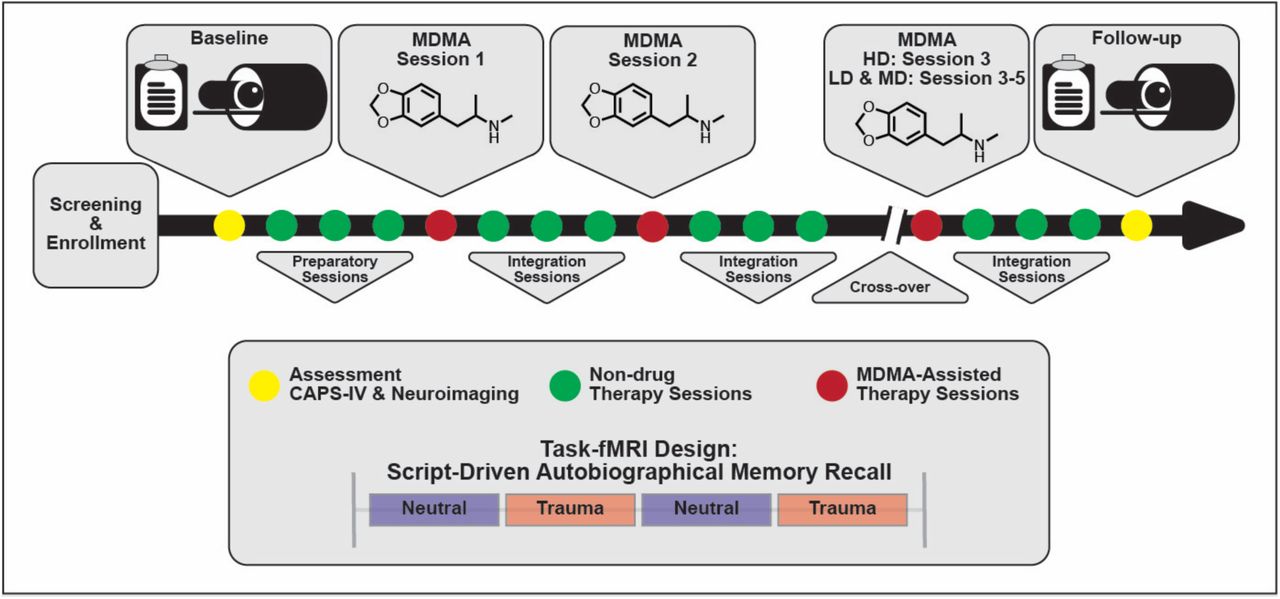
The present study analyzed data from a sub-study (NCT02102802) of a Phase 2 randomized, double-blind, dose-response trial of MDMA-AT in veterans and first responders with severe and chronic PTSD (NCT01211405) (Mithoefer et al. 2018). A detailed study description of the parent study can be found in (Mithoefer et al. 2018), here we summarize the study design. Participants were recruited and screened between November 10, 2010 and January 29, 2015. Veterans and first responders with severe PTSD as measured by a Clinician-Administered PTSD Scale (CAPS-IV) (Blake et al. 1990) total severity score of 50 or more were enrolled in the study and received three 90-minutes preparatory therapy sessions. In Stage 1, participants were randomly assigned to three groups (1:1:2) that received blinded 30, 75 or 125 mg MDMA HCl (followed by a supplemental half-dose unless withheld or declined) with therapy in two 8-hour dosing sessions. Each of the dosing sessions was followed by three non-drug 90-minute follow-up integration sessions that were one month apart. Psychological assessments were collected at baseline and the primary endpoint which was one month after the second dosing session. In Stage 2 of the study, the blind was broken and participants originally in the HD (125 mg) group participated in a final open-label session at the same dose, while participants originally in the LD (30 mg) or MD (75 mg) groups participated in three HD (100 – 125 mg) open-label sessions. Each dosing session in Stage 2 was followed by three non-drug 90-minute integrative sessions. Psychological assessments were collected approximately 2 months and 12 months after the final dosing session in Stage 2. Psychological assessment included the clinician-administered measures CAPS-IV and Global Assessment of Functioning (GAF; general psychological function; Guze 1995), and self-reported measures including Beck Depression Inventory-II (BDI-II; depression symptoms; Beck et al. 1996), Pittsburgh Sleep Quality Index (PSQI; sleep quality; Buysse et al. 1989), Post-Traumatic Growth Inventory (PTGI; perceived growth following trauma; Tedeschi and Calhoun 1996), and the Dissociative Experiences Scale II (DES-II; symptoms of dissociation; Carlson and Putnam 1993).
Participants in the parent study were able to opt into the MRI-based sub-study after which they provided written informed consent approved by the Medical University of South Carolina Institutional Review Board. They were screened for additional neuroimaging related eligibility criteria and were excluded for any conditions that could render MRI unsafe. Following baseline CAPS-IV assessment in the parent study, sub-study participants worked with investigators to create two scripts: one describing a personally traumatic event and one reflecting their typical morning routine at home. Two audio recordings, each six minutes in length, were created from the participant’s reading of each script. Each audio recording was divided into two 3-minute blocks for the task-fMRI. All participants were imaged at baseline, prior to therapy, and again at the follow-up visit two months after their final dosing session. LD (N = 2) and MD (N = 2) participants were additionally imaged after the primary endpoint visit in Stage 1 (one month following their second dosing session), however the small sample sizes prevented any meaningful analysis with these scans. The present analysis focuses on the pre- and post-therapy effects of MDMA-AT on fMRI biomarkers, and thus uses the scans collected at pre-treatment (baseline) and at least 2 months after the largest dose of MDMA (follow-up).
2.2 MRI acquisition
At each scanning session, participants underwent MRI on a 32 channel 3T Siemens system. T1 anatomical scans with TR/TE=1900/2.34 ms and 0.9×0.9×1.0 mm voxel size were collected, followed by two identical task fMRI (design described below) (TR/TE = 2200/35ms, 3.0 mm isotropic voxel size, length of each scan = 14:25 min) and one resting state fMRI (TR/TE = 2000/30ms, 3.3×3.3×3.0 mm voxel size, length = 5:00 min).
Participants’ 6-minute trauma and neutral audio scripts were divided into two three-minute trauma and neutral blocks each (See 2.1 Trial design for description of audio recordings). During fMRI, participants were presented with the visual cue “allow” and instructed to allow themselves to experience the scripts as their audio recordings were played for both neutral and trauma blocks. Each task scan had an alternating block design (neutral 1, trauma 1, neutral 2, trauma 2) with an 18 second ‘rest’ period at the start of the scan and between each block, and about a minute of rest at the end of the scan. The precise length of each audio block was 2.95 min.
2.3 Image preprocessing
FreeSurfer (Dale, Fischl, and Sereno 1999) was applied to the T1s to create white matter (WM), gray matter (GM) and cerebrospinal fluid (CSF) segmentations. FMRIB Software Library (FSL) (Smith et al. 2004) was used for 1) brain extraction, 2) registration between T1s and fMRI’s (brain-boundary registration, non-linear, full-search), 3) high-pass filtering 4) slice-time and 5) motion correction.
2.4 Activation analysis: brain response to trauma versus neutral audio listening
FSL’s FEAT tool (Woolrich et al. 2004) was used for fitting a general linear model (GLM) to the voxelwise timeseries for each task scan after spatial smoothing using a Gaussian kernel function (6 mm full width at half maximum (FWHM)). We constrained our search to four bilateral regions of interest (ROIs) in PTSD symptom provocation (Sartory et al. 2013) using the Desikan-Killiany atlas (Desikan et al. 2006): the precuneus, isthmus cingulate (containing the retrosplenial cortex), rostral anterior cingulate, and amygdala. For 1st-level analysis, models were generated for the neutral block, the trauma block, and a contrast of the two (trauma >b neutral). Confound explanatory variables (EVs) included the temporal derivative of each block, 5 nuisance regressors each for WM and CSF signal, and 6 directions of motion (x/y/z mm/rad). Second-level analysis averaged the models from each of the two task scans performed at each time point. Third-level analyses, using a two-sided, one-sample t-test (FSL randomize; Winkler et al. 2014) identified group-level response for the contrast model (i) at baseline, and (ii) at the two-month follow-up. A final third-level analysis (iii) compared the group-level responses to the contrast model at baseline and follow-up using a two-sample, two-sided, paired t-test (FSL randomize; Winkler et al. 2014). Third-level results were corrected for multiple comparisons using threshold-free cluster enhancement (TFCE; p < 0.05) (Smith and Nichols 2009).
2.5 Functional connectivity
Functional connectivity here is defined as the statistical relationship (in this case Pearson correlation) between two brain regions’ blood-oxygen level dependent (BOLD) time series extracted from fMRI scans.
Prior to extraction of functional connectivity, in addition to the preprocessing steps taken in 2.3, fMRI data were further denoised using an in-house pipeline (https://github.com/kjamison/fmriclean). FMRIs were bandpass filtered and regressed for 24 motion confounds (Friston et al. 1996), 5 nuisance regressors each for WM and CSF, and one for global GM signal. The first five frames (scanner start-up noise) and confound frames were discarded. Gray matter was further parcellated into 68 cortical and 16 subcortical regions of interest (ROIs) (Desikan et al. 2006). The cerebellum was excluded due to incomplete scan coverage of this structure.
In the pre-hoc analysis of the resting-state fMRI data, resting-state functional connectivity (RSFC) between the right and left amygdala and right or left hippocampus was evaluated. One-tailed, paired t-tests were used to compare the 4 RSFC measures before and after MDMA-AT. Functional connectivity during task fMRI between the four regions of interest (bilateral amygdala and hippocampus) and the rest of the brain was assessed. This was done with the time-series extracted from the (a) neutral block and (b) trauma block, separately. See the Supplemental Information for functional connectivity analysis using the entire scan. Group-level changes from pre-to post-therapy in the strength of functional connections were assessed using two-tailed, paired t-tests.
After obtaining whole-brain activation and functional connectivity results from the task data, which highlighted the precuneus as a region of interest, we performed a post-hoc analysis of the RSFC between both amygdalae and hippocampi with the right and left precuneus. We used two-tailed, paired t-tests to compare the 8 RSFC measures before and after MDMA-AT. Pearson correlations were calculated between individuals’ changes in functional connection strength and change in CAPS-IV total severity scores (follow-up – baseline). All p-values were corrected for multiple comparisons using the Benjamini-Hochberg algorithm (Benjamini and Hochberg 1995) where indicated.
3. RESULTS
3.1 CAPS significantly decreases after HD MDMA-AT
Ten participants enrolled in the sub-study, and one withdrew consent after baseline due to anxiety in the MRI scanner, leaving nine participants with MRI data at both time points (6 male, aged 41.3; standard deviation (SD) = ± 10.9 years; 8 veterans and 1 first responder). All participants self-reported moderate depression as indexed by BDI-II (Beck et al. 1996). One subject’s baseline resting-state fMRI was truncated due to technical issues, leaving eight subjects for resting-state analysis and nine for the task fMRI analysis. Mean (SD) CAPS-IV total severity scores of the nine individuals pre-and post-MDMA-AT were 86 (± 16) and 39 (± 25), respectively, representing a significant decrease in PTSD symptom severity between the two time points (Figure 2; N=9, t = 6.36, p = 0.00022). The average percent decrease in CAPS was 57 (± 26)%. Results on all participants enrolled in the Phase 2 parent trial have been previously reported (Mithoefer et al. 2018).
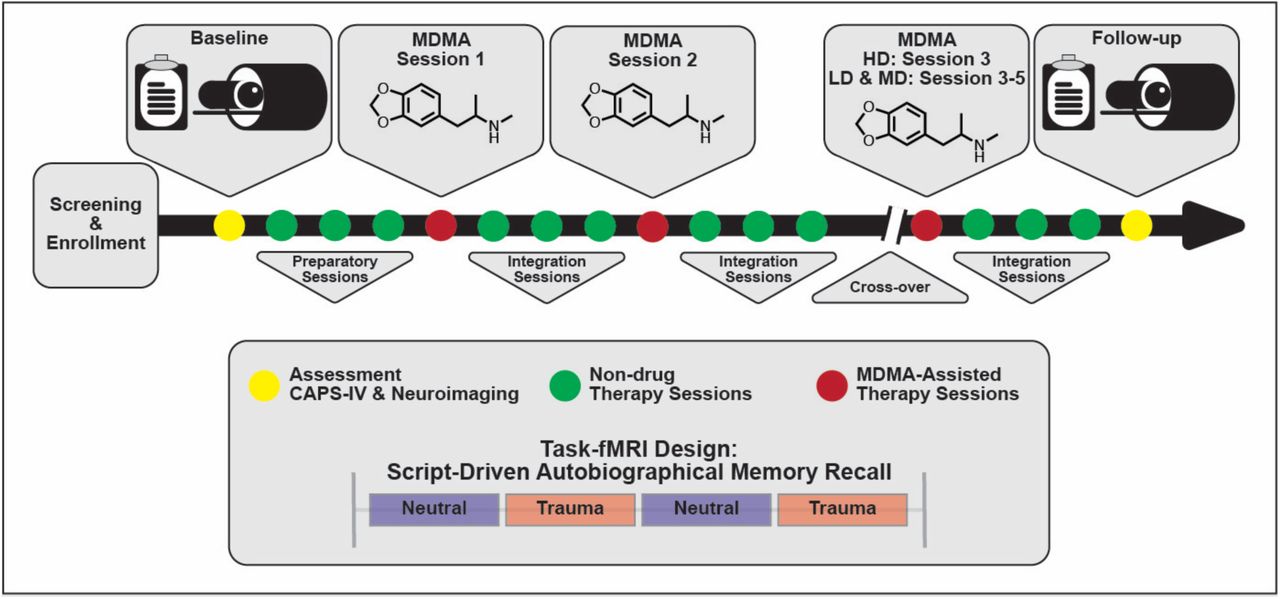
Study design. Subjects were assessed and imaged at the start of the study (baseline). All subjects (low dose (LD), medium dose (MD), and high dose (HD)) underwent three non-drug preparatory therapy sessions prior to their first MDMA dosing session. Each MDMA session was followed by three non-drug integration therapy sessions. After MDMA Session 2 and the subsequent integration sessions, subjects were assessed and the dosing blind was broken. HD subjects completed their final set of drug and non-drug therapy sessions unblinded, and LD/MD subjects crossed over into the HD arm where they completed three sets of drug and non-drug sessions, now with the higher dose and unblinded. All subjects were assessed and underwent MRI approximately two months following their last HD MDMA session.

Patient CAPS-IV total severity scores at the baseline (pre-therapy) and two-month follow-up (post-therapy) scanning sessions. Black and red lines indicate group means and medians, respectively. A significant reduction is PTSD severity was observed after MDMA-AT (baseline > follow-up; N=9, t = 6.36, p = 0.00022).
3.2 Amygdala-hippocampal RSFC increases after HD MDMA-AT
The RSFC was assessed between the amygdala and hippocampus before and after MDMA-AT and the strengths of these connections are illustrated in Figure 3. All connections trended towards increased functional connectivity after therapy compared to before therapy (using a one-sided paired t-test), with left amygdala to both hippocampi having significant increases (t = -2.97, uncorrected p = 0.0104, pFDR = 0.0416 and t = -2.11, uncorrected p = 0.0361, pFDR = 0.0722 for left and right hippocampus, respectively. N=8).

RSFC between the amygdala and hippocampus before and after MDMA-AT. P-values from a one-sided, paired t-test have been corrected for multiple comparisons. Black and red lines indicate group means and medians, respectively. (N = 8; t-statistics indicate baseline > follow-up; * uncorrected p < 0.05, ** pFDR < 0.05).
Individual-level pre-to-post-therapy changes between the strength of these functional connections were then correlated with changes in CAPS scores. All four correlations showed that larger increases in connectivity strength after therapy were associated with larger reductions in symptom severity (improvement in PTSD). Only one of these (right amygdala to left hippocampus) was significant before correction (N = 8; R = -0.773, uncorrected p = 0.0244, pFDR = 0.0976).
3.3 Increased precuneus and retrosplenial cortex activation during symptom provocation
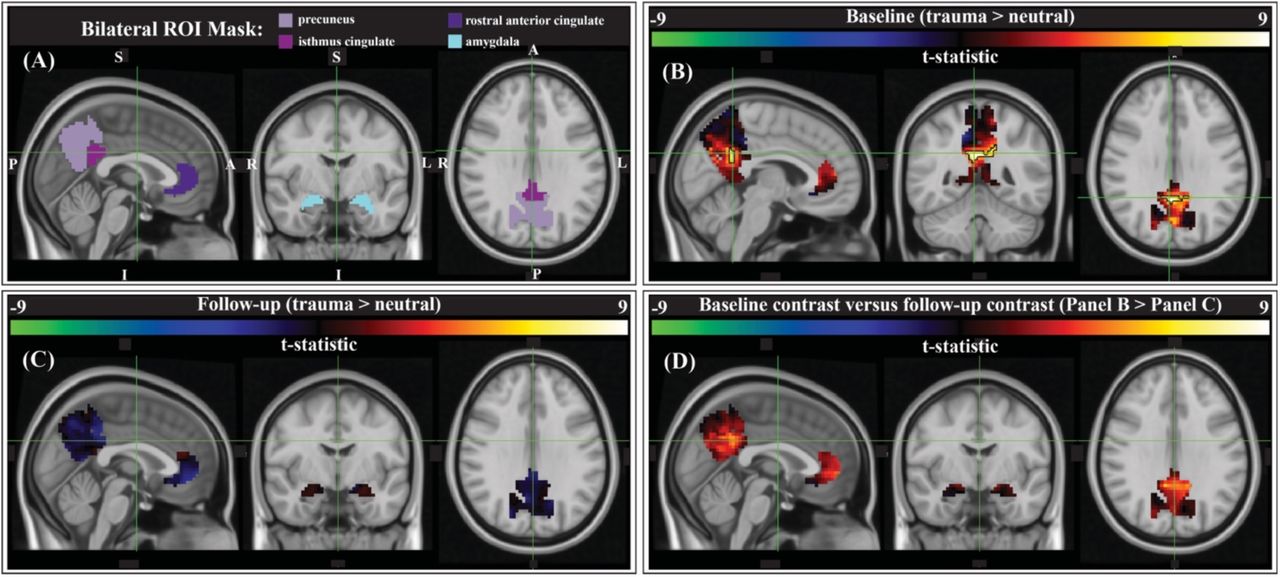
We compared script-driven activations (trauma > neutral) in four pre-determined ROIs: the bilateral precuneus, isthmus cingulate (anatomical gyrus containing the retrosplenial cortex), rostral anterior cingulate cortex (ACC), and amygdala (Figure 4A). These regions have previously been shown to activate more during trauma-related stimuli than neutral stimuli in PTSD patients (Sartory et al. 2013). Before therapy, there tended to be stronger responses to the trauma script versus the neutral script in our ROIs, as evidenced by the generally positive t-statistics (Figure 4B). After correction using threshold free cluster enhancement (TFCE), there was significantly greater activation during the trauma scripts compared to the neutral scripts in a cluster on the border of the bilateral precuneus and bilateral isthmus cingulate/retrosplenial cortex (Figure 4B, black outline; N=9, cluster corrected, p<0.05). After therapy, there was less contrast between the two scripts, and no significant clusters (Figure 4C). Finally, we assessed the differences in the contrast model before and after therapy (baseline > follow-up). There was generally greater (positive) contrast between the trauma and neutral scripts at baseline than at follow-up, however no clusters survived correction for multiple comparisons (Figure 4D).
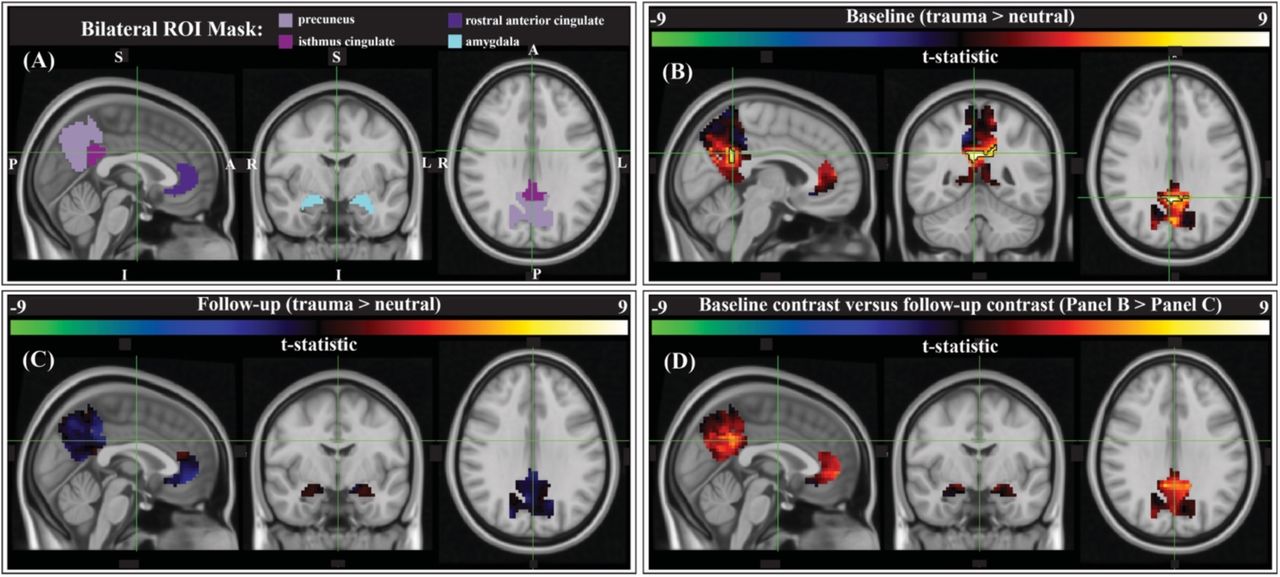
Group-level activation contrasts for trauma versus neutral script listening tasks. (A) Results were constrained to four bilateral anatomical ROIs from PTSD symptom provocation literature (Sartory et al. 2013): the precuneus, isthmus cingulate (retrosplenial cortex), rostral anterior cingulate cortex (ACC), and the amygdala (Desikan et al. 2006). Panels B, C, & D show the t-statistic from the corresponding comparison. (B) Pre-therapy there is a significant cluster within the bilateral precuneus/isthmus cingulate (outlined in black; center of gravity: MNI -1, - 51, 28, 60 voxels (1620 mm3), peak z-value = 2.89; p < 0.05, cluster corrected; N = 9). (C) There were no significant activation contrast clusters after therapy. (D) Comparing the group-level contrasts between time points (baseline > follow-up), there were no clusters that met significance thresholds after correction.
3.4 MDMA-AT associated changes in task functional connectivity
We compared the pre- and post-therapy strength of amygdalae and hippocampal functional connections with the rest of the brain during the neutral and trauma blocks separately (Figure 5A/B). There were several connections with a trend for change from pre-to post-therapy (SI Tables 1 and 2). After correction for multiple comparisons, the left hippocampus and left caudal middle frontal gyrus functional connectivity during trauma recall was significantly reduced at the two-month follow-up post-therapy (N = 9, t = 7.13, pFDR = 0.0082).

Paired t-statistics shown for differences in functional connectivity between all brain regions and the amygdalae and hippocampi during (A) neutral memory audio listening and (B) trauma memory audio listening (N = 9; baseline > follow-up; * two-tailed p < 0.05, uncorrected; ** pFDR < 0.05, corrected).
Individual-level pre-to-post therapy changes in the functional connections calculated in the previous section were then correlated with the individual-level reductions in CAPS scores (Figure 6). Many significant correlations (corrected and uncorrected) were positive, meaning that larger reductions in connectivity from pre-to post-therapy corresponded to larger improvements in PTSD symptomology (SI Tables 3 and 4). Following correction for multiple comparisons, the two positive correlations between changes in right amygdala and bilateral precuneus connectivity during neutral memory recall and CAPS scores remained significant (Figure 6C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pearson correlation values between changes in each functional connection during (A) neutral memory audio listening and (B) trauma memory audio listening and changes in CAPS scores. (C) Scatter plots of the significant correlations from the neutral condition (N = 9; follow-up – baseline; * p-value < 0.05, uncorrected; ** pFDR < 0.05, corrected).
3.4 Post-hoc analysis of amygdala and hippocampus RSFC with the precuneus
As change in the task-based functional connectivity between the amygdala and precuneus was found to be related to improvement of PTSD severity scores, we performed post-hoc analyses of RSFC between the precuneus and amygdalae and hippocampi to identify 1) any changes before and after therapy and 2) any correlations between change in functional connectivity and change in CAPS. We did not find any pre-and post-therapy changes in functional connection strength of amygdalae or hippocampi with the precuneus. Pre-to-post therapy changes in the RSFC between the right amygdala and both the right and left precuneus correlated with the reductions in CAPS scores, albeit at a trend level (N = 9; R = 0.687, uncorrected p = 0.0599; R =0.667, uncorrected p = 0.0709 for the right and left precuneus, respectively), reflecting the task-based functional connectivity finding during the neutral block.
3.5 Correlations with other outcome measures
Although not the primary focus of our analysis, we also repeated the previous correlations using other secondary outcome measures in place of CAPS-IV total severity scores. Namely, changes between baseline and follow-up in the BDI-II (depression symptoms), the PSQI (sleep quality), the PTGI (perceived growth following trauma), the DES-II (symptoms of dissociation), and the GAF (general psychological function) were used. Following correction for multiple comparisons, only three correlations remained significant and, notably, all three were between changes in task functional connectivity and symptoms of dissociation. During the neutral task, reduction in functional connectivity between the left hippocampus and the left pre- and post-central gyrus (motor) at follow-up compared to baseline correlated with reductions in dissociative symptoms (lower DES-II scores; pre-central: R = 0.930, pFDR = 0.0226; post-central: R = 0.904, pFDR = 0.0346). During the trauma task, reduction in functional connectivity between the right amygdala and right insula at follow-up compared to baseline correlated with reduction in dissociative symptoms (R = 0.925, pFDR = 0.029).
DISCUSSION
We report signatures of brain response during rest and audio listening task in eight veterans and one first-responder with clinically diagnosed chronic and severe PTSD before and two-months after MDMA-assisted therapy. We found a significant reduction in CAPS-IV total severity scores after therapy, indicating our sub-study participants mirrored the results from the parent study (Mithoefer et al. 2018). RSFC between the amygdala and hippocampus was significantly strengthened post-therapy, providing more evidence that modulation of amygdalae-hippocampal RSFC may be an important component of MDMA-AT for PTSD (Sripada et al. 2012; Carhart-Harris et al. 2015). We also found participants had increased activation in the precuneus/retrosplenial cortex while listening to traumatic versus neutral memory narrations pre-therapy, and that no significant contrast existed after MDMA-AT (the comparison between the two time points was also not significant). Finally, the pre-to post-therapy reduction in functional connectivity between the right amygdala and right precuneus during neutral memory audio strongly and significantly correlated with PTSD symptom improvement.
Previous work quantifying functional connectivity in PTSD (Sripada et al. 2012) and acute MDMA administration in controls (Carhart-Harris et al. 2015) suggests one mechanism of MDMA-AT may be to increase pathologically lowered amygdala-hippocampal RSFC (Feduccia and Mithoefer 2018). The amygdala is associated with fear expression, threat recognition, and heightened response to emotional memories and is often dysregulated in patients with PTSD (LeDoux 2003; Pitman et al. 2012; Liberzon et al. 1999; Etkin and Wager 2007; Bremner et al. 2005; Harnett, Goodman, and Knight 2020). The hippocampus also plays a central role in PTSD as it is thought to provide contextual information important for cognitive-affect during memory recollection (Harnett, Goodman, and Knight 2020; Pitman et al. 2012). Sripada et al (2012) found combat veterans with PTSD had reduced amygdala-hippocampal RSFC compared to combat-exposed controls, leading them to speculate that this may relate to an inability to contextualize affective information in PTSD. Carhart-Harris et al (2015) demonstrated that amygdala-hippocampal RSFC is increased acutely in MDMA administration compared to placebo and this increase occurred in a manner that correlated with the drug’s subjective effects at a near-significant level, leading these researchers to propose that this functional connection was a primary target of MDMA-AT. Prior to our analysis (although after the study was designed and the data collected), we hypothesized that the RSFC between the amygdala and hippocampus would be higher after MDMA-AT compared to pre-therapy levels, which we confirmed. We also found that the amount of increased amygdala-hippocampal RSFC after MDMA-AT positively correlated with PTSD symptom improvement at a near-significant level. Our current findings provide further evidence that one mechanism driving MDMA-AT’s treatment of PTSD could be an increase in amygdala-hippocampus RSFC.
We next sought to study brain-response during autobiographical memory listening to draw additional conclusions about MDMA-AT’s effects in individuals with PTSD. Before therapy, participants had larger activation in a cluster of voxels located in the bilateral precuneus and retrosplenial cortex during an individualized trauma script listening task compared to neutral audio script listening. The precuneus is involved in self-processing operations (e.g. first-person perspective taking), episodic memory retrieval, and visual-spatial imagery (Cavanna and Trimble 2006). Its activation is often correlated with activation of the retrosplenial cortex (Cauda et al. 2010), which is implicated in autobiographical memory recollection (Svoboda, McKinnon, and Levine 2006; Spreng, Mar, and Kim 2009). The retrosplenial cortex is also found to be consistently activated by emotionally salient stimuli, and has been proposed to play a role in the interaction between emotion and memory (Maddock 1999). We conjecture that increased activation in these regions during traumatic compared to neutral audio listening (Figure 4B) could be related to an increased intensity of the recollection or re-experiencing of traumatic memories compared to neutral ones for patients before therapy. At 2-month follow-up to MDMA-AT, there was no significant difference in the trauma vs neutral script activation of this area (Figure 4C). Although the longitudinal comparison of these two time points also showed no significant difference after correction for multiple comparisons, the positive t-statistics indicate that the contrast between trauma and neutral was generally larger at baseline, particularly in the precuneus/retrosplenial cortex and the anterior cingulate cortex (ACC) (Figure 4D). Larger studies may allow more statistical power to identify longitudinal differences if they do indeed exist. These findings are consistent with previous studies — a meta-analysis by Sartory et al (2013) (the source of our ROIs) found that PTSD patients had greater activations in the precuneus and retrosplenial cortex, as well as the bi-lateral amygdala and anterior cingulate cortex during trauma-related stimuli compared to neutral stimuli. In addition, other longitudinal studies of individuals with PTSD have found the amount of decrease in precuneus/posterior cingulate cortex (PCC; of which the isthmus cingulate/retrosplenial cortex is a part) activation during symptom provocation at the end of study to be correlated with amount of reduction in symptom severity (Garrett et al. 2019; Ke et al. 2016).
PTSD is often associated with hyperactivity in the amygdala (Pitman et al. 2012); the acute administration of MDMA in healthy volunteers decreases blood flow to the amygdala during rest (Carhart-Harris et al. 2015) and attenuates its response to angry faces (Bedi et al. 2009). We had hypothesized that we would observe hyperactivity of the amygdala to trauma versus neutral scripts at baseline and that MDMA-AT would attenuate this response, however we observed neither. It is important to note inconsistencies in the literature here. Amygdala hyperactivity in PTSD is not always observed, possibly due to differences in subtypes, sex, cultural representations, or choice of paradigm (van Huijstee and Vermetten 2018; Lanius et al. 2001; Lanius et al. 2002; Helpman et al. 2021; Chiao et al. 2008; Liddell and Jobson 2016). Additionally, while MDMA did suppress amygdala activity during rest and in response to angry faces as previously mentioned, there was no observed impact on its response to autobiographical memories (Carhart-Harris et al. 2014). While activation-based analyses deserve continued attention in future studies to rectify these inconsistencies, functional connectivity is a complimentary approach we can use to extract additional information from fMRI.
In a task-free setting, amygdala activity has been found to be anti-correlated with activity in areas involved in effortful regulation of affect, including the precuneus (Roy et al. 2009; Zhang and Li 2012). In healthy adults, amygdala-precuneus functional connectivity increases following the acute exposure to social stress (Veer et al. 2011, 2). This functional connection has been implicated in mood disorders including depression (Cullen et al. 2014; K. D. Young et al. 2018; Wei et al. 2018; Feurer et al. 2021; Peng et al. 2020), bipolar disorder (Stoddard et al. 2015; Singh et al. 2015) and anxiety (Strawn et al. 2012; Toazza et al. 2016), childhood emotional maltreatment (Werff et al. 2013), and PTSD (Nicholson et al. 2015; Bluhm et al. 2009; R. A. Lanius et al. 2010). One study found adults who experienced childhood emotional maltreatment show increased (decreased magnitude negative) connectivity between the amygdala and precuneus compared to controls (Werff et al. 2013), and another found PTSD patients with dissociative sub-type had increased connectivity compared to PTSD patients without dissociative sub-type (Nicholson et al. 2015). Here, we find a strong, significant correlation between pre-to post-therapy decreases in right amygdala-precuneus functional connectivity during neutral scripts and pre-to post-therapy decreases in CAPS-IV. Attenuated correlation between the activity patterns of these two regions at follow-up possibly suggests a decreased intensity of recalled events, less ‘re-experiencing’, or reduced negative affect.
It is worth noting here that increased amygdala-precuneus functional connectivity during both rest and positive autobiographical memory recall was found to correlate with improved depressive symptoms in adults using real-time fMRI neurofeedback targeting amygdala activation (K. D. Young et al. 2018). Depression is characterized by decreased arousal (depressed mood) and diminished positive affect, therefore these findings may be reflective of an increased emotional response to positive memories that is associated with symptom improvement (Dunn et al. 2020). However, another study in adolescents found that those with depression had greater amygdala-precuneus RSFC compared to healthy controls (Cullen et al. 2014), so replication is necessary. It seems likely that this functional connection is relevant to cognitive affect in mood disorders, however more research is needed to determine if these findings are reflective of differences in depression and PTSD, or the context in which the study populations were being imaged (i.e. studying processes and tasks related positive affect in the case of depression and negative affect in the case of PTSD). Considering the co-morbidity of depression and PTSD (Brady et al. 2000), amygdala-precuneus functional connectivity may deserve increased attention in future studies.
Also of interest were the significant correlations found between reduction in symptoms of dissociation and changes in task functional connectivity. During neutral memory audio listening, the reduction of left hippocampus to both left motor (pre- and post-central) gyri functional connectivity correlated with reduction in dissociative symptoms. These two functional connections appear to be higher in cases of trait anxiety (Yang et al. 2017). Lastly, MDMA acutely alters insula functional connectivity (Walpola et al. 2017), a region implicated in anxiety (Etkin and Wager 2007). Baur et al. (2013) provided evidence that amygdala-insula functional connectivity indexes individual differences in state anxiety. Here, we find that reductions in right amygdala-insula functional connectivity during trauma audio listening after MDMA-AT correlates with reductions in dissociative symptoms, possibly relating to reduced anxiety or dissociation during traumatic memory recall after treatment.
PTSD is characterized by decreased fear extinction in response to trauma-related stimuli. One possible mechanism through which MDMA-AT operates is enhanced reconsolidation and/or fear extinction processes (Feduccia and Mithoefer 2018). Several studies with MDMA implicate reconsolidation or fear extinction processes, and while it is currently unclear whether MDMA acts on only one or both, it is important to note that the two interact (Suzuki et al. 2004). Rodent models have demonstrated that the administration of MDMA prior to extinction learning enhances post-acute extinction retention and this effect is blocked by acute and chronic treatment with a serotonin transporter inhibitor (M. B. Young et al. 2015; B. Young et al. 2017). Hake et al. (2019) found that MDMA administered during extinction learning phases did not enhance fear extinction memory, while MDMA administration during reconsolidation phases resulted in prolonged reductions in conditioned fear. In addition, MDMA administered prior to trauma-cue exposure (reconsolidation phase) in rodents resulted in reduced stress-related behavioral responses 7 days later (Arluk et al. 2022). A randomized, controlled clinical trial in healthy humans found that those who were administered MDMA (100 mg) prior to extinction learning showed a higher rate of total extinction retention 48 hours later compared to the placebo group (Maples-Keller et al. 2022). Doss et al. (2018) found that 1 mg/kg of MDMA in healthy humans attenuated the encoding and retrieval of salient details from positive and negative stimuli (but not neutral stimuli), suggesting an ability for MDMA to alter emotional memory representation. Interestingly, a fMRI study in healthy humans found decreased activation in the precuneus/PCC during fear extinction learning (Ridderbusch et al. 2021), regions highlighted by our present study and others in PTSD (Garrett et al. 2019; Ke et al. 2016).
LIMITATIONS
The small sample size of the present study and the lack of a control population (e.g. trauma-exposed controls) may decrease the generalizability of these findings. The trial design was placebo-controlled for dose-response (low, medium, and high), however, the follow-up scans used in this study were after the breaking of the blind and dose cross-over (low/medium to high) had occurred. For neuroimaging studies, comparisons with control populations are helpful for contextualizing longitudinal changes in brain response and provide information about whether changes in patient populations represent an abnormal response being restored to normality or a compensatory mechanism. In addition, multi-point imaging of healthy control or non-treatment (placebo) groups allow for the quantification of test-retest variability.
Here it must be discussed that PTSD is a disorder exhibiting at least two major sub-types with characteristically opposing phenomenological and physiological responses to symptom provocation, which may explain inconsistencies in the PTSD neurobiology literature (van Huijstee and Vermetten 2018). In addition to sub-type heterogeneity, males and females with PTSD may also differ in their neural responses (Helpman et al. 2021). Limited by our sample size, we did not investigate these sub-groups in this study.
The accepted standard for assessing PTSD severity is the Clinician-Administered PTSD Scale (Blake et al. 1990). Specifically, CAPS-IV was used in this study. CAPS-IV involves an hour-long semi-structured interview with a clinician and, though comprehensive, faces limitations. In their baseline CAPS-IV assessment, and subsequently thereafter, patients were asked to refer to an index trauma that was measured throughout the study. This may present an issue in accurately assessing global PTSD severity if an adjacent or un-related trauma surfaces during therapy and becomes the prominent driver of their symptoms. These issues, combined with difficulty in blinding and expectancy effects, present additional challenges in accurately mapping fMRI metrics to clinical outcomes. At the same time, these challenges motivate the need to improve our ability to detect biomarkers of PTSD to accompany psychological indexes.
The task design used in this study examined differences in brain response to personalized audio scripts generated from narrations of traumatic and mundane (neutral) memories. Many different stimuli have been used in fMRI studies of PTSD (R. A. Lanius et al. 2006; Sartory et al. 2013; Patel et al. 2012), each providing its own unique advantages and disadvantages. Our present design optimizes personal relevance of the stimuli; however, this has the consequence of presenting each subject with a different set of stimuli, whereby brain responses within each block are not time-locked across participants. Also, it has previously been shown that PTSD survivors take longer to retrieve unrelated autobiographical information when listening to taped imagery scripts of their traumatic memories (Kleim, Wallott, and Ehlers 2008). This suggests the possibility that those with the most severe PTSD will take the longest to cognitively transition to the neutral block from the trauma block. If this is true, then there would perhaps be an inverse-“U” relationship between PTSD severity and contrast between the trauma and neutral conditions, if the blocks are not spaced far enough apart to allow adequate time for patients to return to a baseline level of cognitive functioning.
Finally, the pre-specified aim of this study was to estimate longitudinal (baseline to 2-months after final MDMA session) changes in ROI response to traumatic audio scripts. Between the start of data collection and analysis, new literature emerged (Carhart-Harris et al. 2015) implicating amygdala-hippocampus RSFC as a potential target of MDMA-AT, compelling us to expand our analysis beyond the pre-specified aims.
CONCLUSION
We report functional brain changes associated with MDMA-AT in participants with PTSD. We provide further evidence that MDMA-AT may act through strengthening the RSFC between the amygdala and hippocampus, a connection which is weaker in PTSD populations (Sripada et al. 2012) and increased acutely by MDMA in healthy volunteers (Carhart-Harris et al. 2015). We also provide preliminary evidence that MDMA-AT alters amygdala and precuneus response during symptom provocation. These regions, associated with fear response and self-referential processing, respectively, are commonly found to be hyperactive in PTSD patients (Sartory et al. 2013; Patel et al. 2012). However, the reduction of right amygdala-precuneus functional connectivity co-varying with symptom reduction in PTSD is a novel finding. More research is necessary to confirm this result and determine if this is an effect specific to MDMA-AT, or if it would be found in other longitudinal treatment cohorts as well.
Data Availability
All data in the present study will be made available upon reasonable request to the authors following peer-review and publication.
Acknowledgements
The authors would like to thank the candidates who were willing to be screened for eligibility; the participants in the trial who contributed their data; S. Sadler for her dedication as the Study Coordinator; S. Braswell for serving as night attendant, M. Wagner and J. Wymer for Independent Rater assessments. The authors would also like to thank: J. Holland who supported this trial as a Medical Monitor; L. Jerome for her numerous and varied contributions to this Clinical Development Program since prior to its inception with global systematic literature reviews and medical coding; C. Hennigan for data management; R. Matthews and B. Shechet for clinical operations and monitoring; J. Sonstroem and A. Seltzer for randomization support and system programming; E. Sola, Y. Gelfand, and B. Cohen for conducting adherence ratings to facilitate standardization of therapy; I. Gorman for development of adherence ratings; A. Wilens for supporting video recording. The authors would like to thank Dr. Edmund Higgins for catalyzing the first discussions about acquiring imaging data in the MAPS trial.
Footnotes
Role of Funding Sources: The clinical trial was sponsored by the Multidisciplinary Association for Psychedelic Studies (MAPS), a 501(c)(3) nonprofit organization. MAPS provided the MDMA and fully funded this study from private and foundation donations. MAPS Public Benefit Corporation (MAPS PBC), wholly owned by MAPS, was the trial organizer. SPS is supported by the National Science Foundation Graduate Research Fellowship (Grant No. DGE-1650441). AK is supported by the National Institutes of Health (RF1MH123232 and R01NS102646).
Conflict of Interest: MM was paid as a contractor by MAPS PBC. AE, CP, AC, and JW received salary support for full-time employment with MAPS PBC. BYK and RD received salary support for full-time employment with MAPS. MM is on the Clinical Advisory Board of Awakn Life Sciences.
REFERENCES




