
ABSTRACT
Aortic dimensions and distensibility are key risk factors for aortic aneurysms and dissections, as well as for other cardiovascular and cerebrovascular diseases. We tested genome-wide associations of ascending and descending aortic distensibility and area derived from cardiac magnetic resonance imaging (MRI) data of up to 32,590 Caucasian individuals in UK Biobank. We identified 102 loci (including 31 novel associations) tagging genes related to cardiovascular development, extracellular matrix production, smooth muscle cell contraction and heritable aortic diseases. Functional analyses highlighted four signalling pathways associated with aortic distensibility (TGF-β, IGF, VEGF and PDGF). We identified distinct sex-specific associations with aortic traits. We developed co-expression networks associated with aortic traits and applied phenome-wide Mendelian randomization (MR-PheWAS), generating evidence for a causal role for aortic distensibility in development of aortic aneurysms. Multivariable MR suggested a causal relationship between aortic distensibility and cerebral white matter hyperintensities, mechanistically linking aortic traits and brain small vessel disease.
INTRODUCTION
The aorta acts as both conduit and buffer1, conveying oxygenated blood from the heart to the systemic circulation, and dampening the pulse pressure to which peripheral circulations are subjected. Diseases affecting the aorta are common and their complications are associated with a high mortality even in young people. Quantitative aortic traits (aortic dimensions and functional measures) can predict progression of these aortopathies. For example, the elastic function of the thoracic aorta and aortic dimensions are key determinants of rates of growth of thoracic aortic aneurysms2-4. At a population level, aortic traits are also clinically important predictors of risks of cardiovascular and cerebrovascular diseases5-10.
The decline in elastic function with age may be measured as a decrease in distensibility, a factor independently predictive of cerebral microvascular disease, the development of age-related dementia and neurodegenerative changes of Alzheimer’s Disease (AD)11-13. Recent data also have provided evidence for an association of aortic distensibility with cognitive performance in the general population14. White matter hyperintensities (WMH) represent the most common brain imaging feature of small vessel disease and predict mortality and morbidity with aging (including risks of stroke (ischemic and hemorrhagic), dementia, and functional impairment15-18). Aortic stiffness is a stronger predictor of WMH volume than blood pressure or hypertension alone19, 20 and has effects additive to those of hypertension in predicting WMH19-21. The genomic bases of these relationships have not been well explored to date.
With age, aortic stiffening arises from changes in composition of the aortic wall, including degradation of the elastic fibres and decreased cellularity, along with a relative increase in the collagen content of the aorta (although the absolute amount decreases)22, 23. In addition, extracellular matrix proteins themselves undergo conformational and biochemical changes which alter their passive mechanical properties. These remodelling processes are driven by TGF-β signalling pathways and accelerated by oxidative stress and inflammation24 acting on the cells in the aortic wall. These cellular and molecular drivers of worsening aortic elastic function are reflected in macroscopic changes with age.
Here, we used convolutional neural networks for automated aortic segmentation25 to measure ascending and descending aortic areas and distensibilities on cardiac magnetic resonance (MRI) images from UK Biobank, which is currently the largest cardiac imaging epidemiological study26. We have described our approach to derivation of imaging derived quantitative aortic traits and the distribution of these traits in a smaller group from the same population in an earlier report25, 27. We derived six aortic traits (ascending aortic distensibility (AAdis), descending aortic distensibility (DAdis), maximum ascending aortic area (AAmax), minimum ascending aortic area (AAmin), maximum descending aortic area (DAmax) and minimum descending aortic area (DAmin)) in up to 32,590 (depending on the specific trait) UK Biobank participants, who were free from known aortic disease We then performed a genome-wide association study (GWAS) of the six CMR-derived aortic traits and carried out functional analyses and a series of Mendelian Randomization studies to investigate possible causal associations of the aortic measures with aortic aneurysms and brain small vessel disease. We also explored the bidirectional relationship of aortic traits with indices of blood pressure.
RESULTS
Study cohort exclusions are presented in Supplementary Figure S1. The distributions of the aortic traits are shown in Supplementary Figure S2.
SNP-based heritability
We estimated the proportion of the variability in aortic traits that could be attributed to common genetic variation from an analysis of SNP-based heritability (h2SNP) using linkage disequilibrium score regression (LDSC). h2SNP estimates ranged from 0.10 (for DAdis single trait) to 0.41 (for AAmax). We also tested for heritability of distensibility traits using multi-trait analysis (MTAG, h2SNP=0.21 for DAdis and h2SNP=0.24 for AAdis).
Phenotypic and genotypic correlations between traits
We found strong phenotypic and genotypic correlations between maximum and minimum aortic areas (phenotypic r=0.99, p<2.2×10−16; genotypic rg=0.99, p<1×10−50 for the ascending aorta; phenotypic r=0.98, p<2.2×10−16; genotypic rg =0.99, p<1×10−50 for the descending aorta).There were lower correlations between ascending and descending aortic traits (phenotypic r=0.60, p<2.2×10−16 and genotypic rg=0.45, p<1.7×10−25 for the minimum aortic areas and phenotypic r=0.74, p<2.2×10−16 and genotypic rg=0.45, p<9.25×10−7 for distensibilities) consistent with known biological and functional differences along the course of the aorta1. Correlations are presented in Supplementary Figures S3 and S4.
Single and multi-trait aortic GWAS
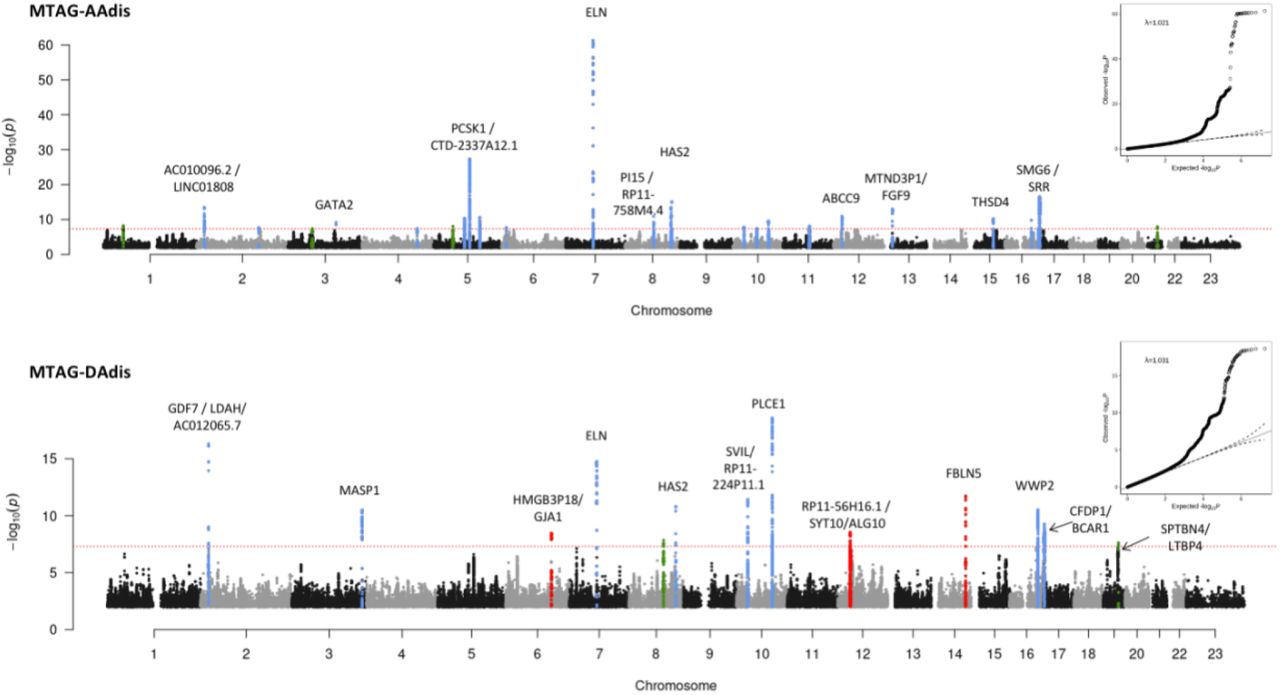
Our stage 1 GWAS (N=32,590 for areas and N=29,895 for distensibility) identified a total of 95 significant loci (using a genome-wide significance threshold of p<5×10−8) across the six traits, 94 of which are autosomal with one localised to the X chromosome. Genomic inflation was within acceptable limits for all traits (λ=1.147 for the area traits; λ=1.047 for the distensibility traits). We took advantage of the correlation between the aortic traits to enhance power for discovery of loci by performing multi-trait analysis (MTAG)28 as a second stage of our GWAS. Use of MTAG combining all six phenotypes increased the number of significant loci for the distensibility traits from 10 to 26 for the ascending aorta and from 7 to 13 for the descending aorta (Table 1), and the total number of significant loci across all aortic traits to 102. Figure 1A shows the Manhattan and QQ plots from GWAS of ascending and descending aortic minimum areas, which overlap almost completely with the findings for the corresponding maximum areas. Figure 1B shows the results of the MTAG analysis of ascending and descending aortic distensibilities. GWAS summary statistics from 9,753,033 variants with a minor allele frequency (MAF) ≥ 0.01 for the stage 1 and stage 2 (MTAG) analyses are shown in Supplementary Figures S5-S8.
Summary statistics are shown for lead SNPs which were genome-wide significant (p<5×10−8) in MTAG analysis (loci in black) apart from the four loci (in green and blue) which were significant in single trait analysis only. Where loci overlapped, different lead SNPs are listed separately under the same locus number. Green = genome-wide significant in only the single trait analysis, not the multi-trait analysis, but also significantly associated with aortic area traits. MAF=Minor Allele Frequency; Chr=chromosome, BP= position (GRCh37). Blue = genome-wide significant in only the single trait analysis of distensibility and in no other aortic traits. *=novel locus not identified in previous GWAS of ascending or descending aortic traits.

Red dashed line shows the genome-wide significance threshold of P = 5 × 10−8. Genomic inflation (λ) = 1.147 (AAmin and DAmin). Annotations of selected loci show the nearest gene and additional manual annotation of likely candidate gene(s) at the locus where appropriate. Blue: locus is genome-wide significant in multiple aortic traits, green: locus is genome wide significant only in the corresponding trait and with nominal significance (p<0.01) in other traits, red: genome-wide significant only in the corresponding trait without even nominal significance in other traits. QQ plots are shown as inserts in corresponding panels.

All six traits were used for the MTAG analysis. Twenty-six association signals were identified in multi-trait analysis of AAdis and thirteen in multi-trait analysis of DAdis. Genomic inflation (λ) = 1.021 (AAdis) and 1.031 (DAdis). Red dashed line shows the genome-wide significance threshold of P = 5 × 10−8. Blue: locus is genome-wide significant in multiple aortic traits, green: locus is genome wide significant only in the corresponding trait and with nominal significance (p<0.01) in other traits, red: genome-wide significant only in the corresponding trait without even nominal significance in other traits. QQ plots are shown as inserts in corresponding panels.
Individual loci were annotated with cis-expression quantitative trait loci (eQTL) and splice quantitative trait loci (sQTL) data from GTEx v829. Twenty-four of the 38 loci associated with distensibilities had lead SNPs which were significant eQTLs or sQTLs for nearby genes in arterial tissue.
The most significant associations with ascending aortic distensibility were: rs7795735, 12.6 kilobases upstream of ELN, a gene encoding elastin; rs201281936, which is in a lncRNA (CTD-2337A12.1) 119 kilobases 3’ of PCSK1 (Proprotein Convertase Subtilisin/Kexin Type 1); rs57130712, which is in a locus spanning SMG6 (SMG6 Nonsense Mediated MRNA Decay Factor) and SRR (Serine Racemase), and which is the most significant eQTL for SRR in arterial tissue (p=2×10−20, normalised effect size (NES)= -0.39)29; and rs34557926, an intronic variant in HAS2 (Hyaluronan Synthase 2).
The strongest associations with descending aortic distensibility were different. The most significant association was with rs61886305, an intronic variant in PLCE1 (Phospholipase C Epsilon 1). This lead SNP is a strong eQTL for PLCE1 in arterial tissue (p=1.1×10−8, NES= -0.20)29. The next strongest association was with rs9306895, an intronic variant in GDF7 (Growth Differentiation Factor 7), which is a strong eQTL for both GDF7 and LDAH (Lipid Droplet Associated Hydrolase) in aorta29 (GDF7: p=3.6 ×10−9, NES= 0.20; LDAH: p=7.1×10−28, NES= 0.53). A locus spanning ELN was associated with DAdis, with the lead SNP less than 1.8kb away from the lead SNP for AAdis at this locus, and in strong LD with it (R2=0.96; D’=1). A locus in FBLN5 (lead SNP rs8014161) was associated with descending, but not ascending distensibility.
There were four loci associated at genome-wide significance with aortic distensibilities which lost genome-wide significance in the MTAG analysis (three associated with AAdis and one with DAdis; see Table 1 for details). Two of these were not significantly associated with any other aortic traits: rs6724315 in PRKCE and rs10857614 in ARHGAP22. The latter is a strong eQTL for ARHGAP22 in aorta (p=2.6×10−46, NES= 0.58), providing additional evidence for its biological relevance.
We compared our association results for aortic areas with those reported for aortic diameters in recent preprints30, 31 based on the same UK Biobank imaging dataset, but using different methods and metrics for aortic dimensions. We replicated 73 of the previously reported genome-wide significant association loci and added associations for two clinically relevant phenotypes (ascending and descending aortic distensibility) to identify a further 31 novel associations. Only four of the previously reported SNPs did not replicate. Inspection of the loci for SNPs that were significant in the analysis by Pirruccello et al30, but not in our own, generally showed SNP p-value signals near the genome-wide significance threshold (p<5×10−8) in our analysis. The small differences between associations in the two studies could arise from differences in the methods used to generate quantitative phenotypes or from the differences in participant exclusions between the two studies (for example, we excluded data from participants with known diagnoses of aortic disease and those who were extreme phenotypic outliers).
Novel aortic loci associated with AAdis included lead SNPs rs7638565 near ADAMTS9, which is a significant eQTL for this gene in aorta (p=5.7×10−7, NES= -0.22) and rs835341, intronic in GPX7 and a strong eQTL in the aorta (p=3×10−95, NES= -0.92). A novel locus associated with descending aortic distensibility included lead SNP rs112009052, an sQTL for LTBP4 in fibroblasts (p=9.6×10−67, NES= 2.9) but not in aortic tissues that has relevance here due to this gene’s role in the TGF-β pathway.
We identified 21 novel associations with aortic areas including lead SNPs in KALRN and COL21A1 associated with ascending aortic areas and SNPs at loci tagging AFAP1, FGF5/BMP3, NOX4, FES and GATA5/LAMA5 associated with descending aortic areas (see Supplementary Figures S5).
Sex-specific aortic trait GWAS analyses
We undertook sex-specific GWAS analysis of the area phenotypes. We did not perform these analyses for distensibility phenotypes due to lack of power with the smaller cohort sizes. We contrasted associations discovered for the men and women (numbers of whom were well-balanced in the cohort) using a z test. There were 18 loci (ten for AAmin and eight for DAmin) at which the differences between sexes were significant (adjusted p <0.05). Seven of these associated loci were not significant even at p<0.05 for one sex, despite reaching genome-wide significance (p<5×10−8) for the other (see Table 2). Amongst these sex-specific loci were rs28699256, a missense variant in in ADAMTS7 associated with ascending aortic area in females, but not males (see Table 2; z-test for sex difference, p=7.6×10−4). This variant in ADAMTS7 is in strong LD with the lead SNP associated with AAmin in the full cohort at genome-wide significance (rs7182642; R2=0.76, D’=0.94). Amongst the other sex-specific signals were four others with functional data supporting potential biological roles in the aorta, all of which were significant only in males: rs72765298 in SCAI, a strong eQTL in aorta (see Table 2; p value for eQTL in aorta=8.9×10−18, NES= -0.48); rs632650, a significant eQTL for ALDH2 in aorta (see Table 2; p value for eQTL in aorta=3.3×10−12, NES= 0.26); rs6573268, associated with DAdis, in CCDC175 which is a significant eQTL for this gene and others in the aorta (see Table 2; p value for eQTLs in aorta: for CCDC175 p=1.2×10−6, NES = 0.35; for RTN1 p=2.4×10−7, NES= 0.33 and for L3HYPDH, p=1.1×10−4, NES= 0.25); and rs35346340 in FES, a strong eQTL for this gene in aorta (see Table 2; p value for eQTL in aorta 2.4 × 10−15, NES= 0.3).
SNPs are shown if they reach genome-wide significance (p<5×10−8) in one sex and are not significant (p>0.05) in the other. Gene = nearest gene. Trait = aortic trait with which association in one sex is genome-wide significant, pval = p value from sex-specific GWAS, beta= effect size from sex-specific GWAS, SE= standard error from sex-specific GWAS, sig = which sex the SNP has reached genome wide significance in, z stat = z statistic for comparison between sexes, z.pval= p value of sex comparison.
An association with rs12663193 (intronic in ESR1) reached genome-wide significance only in females. However, the sex difference was not significant (z-test p value = 0.06).
Gene-based analysis and tissue specificity
We prioritized potentially causal genes at significant loci using two complementary strategies: FUMA32, which integrates positional mapping, eQTL associations and HiC-derived 3D chromatin interactions (see Methods) and MAGMA33, which aggregates SNP associations within genes. In total, FUMA identified 973 candidate genes across the six phenotypes, including 390 protein-coding genes, 164 pseudogenes, 129 lincRNAs, 115 antisense RNAs and 46 miRNAs (Supplementary Figure S9). MAGMA identified 391 candidate genes with an FDR < 0.01 (Supplementary Figure S10).The most significant gene associations (MAGMA) for ascending and descending aortic distensibilities are shown in Table 3.
Most significant 30 genes associated with ascending and descending aortic distensibility (MAGMA). P values are converted to adjusted p-values (FDRs) using the Benjamini-Hochberg procedure.
Four genes (MASP1, PI15, PLCE, TBC1D12 [the last likely tagging the PLCE1 locus]) reached significance for all 6 aortic traits, with ELN at genome-wide significance in all traits except for DAmax, where it was just below the genome-wide significance threshold.
Tissue specificity analysis in MAGMA for genes associated with each phenotype demonstrated that these were significantly enriched for expression in aorta and in coronary artery (p value for enrichment < 1×10−3 in all traits), supporting the validity of our results. See Supplementary Figure S11 for further details.
Gene set enrichment and pathways analyses
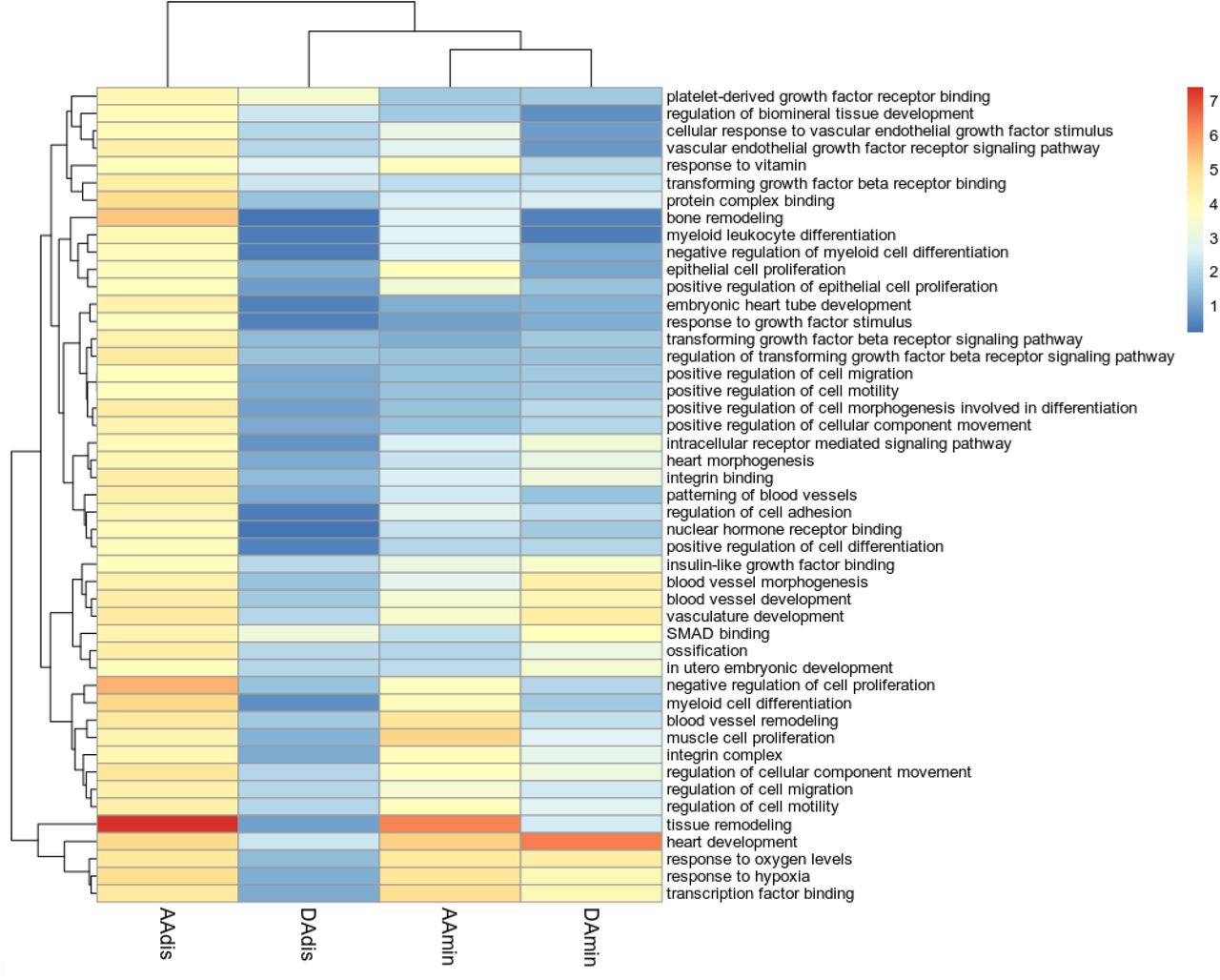
The GO terms identified by MAGMA (Supplementary Figure S12) that were most significantly associated with our aortic phenotypes highlighted processes important for the development of aortic aneurysms and dissection, such as “extracellular matrix structural constituent” and “smooth muscle contraction”, as well as GO terms related to cardiovascular development.
DEPICT implicated similar ontologies and identified three molecular pathways significantly enriched in our data (FDR<0.01) for at least one aortic trait (see Figure 2) and of nominal significance in all other traits: regulation of TGF-β signalling (AAdis nominal p value=2.76×10−5; FDR <0.01), IGF binding (AAdis nominal p value 1.47×10−4; FDR <0.01) and PDGF binding (AAdis nominal p value 7.19×10−5; FDR <0.01). VEGF signalling was significantly associated with ascending aortic distensibility (AAdis nominal p value 5.15×10−5 ; FDR <0.01).

Colour scale denotes the significance of enrichment, -log10 scale. Only GO terms significantly enriched in association with AAdis (FDR<0.01) are presented.
Co-expression network analyses
Using expression data from single cell transcriptomics of the primate aorta34, we generated co-expression modules for aortic endothelial and aortic smooth muscle cells. Using our MAGMA (adj. p-value <0.01) and FUMA gene-based associations (see Methods), we generated functional sub-networks for each aortic trait, highly enriched for our significant genes and identified hub genes for modules expressed in aortic endothelial cells and aortic smooth muscle (see Figure 3 and Supplementary Figures S13-S16). These hub genes include genes involved in smooth muscle cell contraction and differentiation (e.g., ACTB, MYH10, MYL9, NEXN, ARID5B and SVIL), as well as others associated with TGF-β signalling. In endothelial cells, hub genes identified included EDN1 and GATA2.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Co-expression networks for aortic distensibility GWAS genes generated with primate single cell expression data for the aorta34. The co-expression networks were derived from extended models (r>0.2) in aortic endothelial (AoECs) and aortic smooth muscle cells (AoSMCs). Round circles represent genes which were significantly associated with an aortic trait in the current GWAS. Diamonds represent other genes significantly co-expressed in the published single cell data for the cell-type indicated. The deeper the shade of red, the higher the level of expression of that gene in the specified cell-type. The strength of co-expression is denoted by the colour of the lines joining genes with higher correlations indicated by darker lines. “Hub genes” are found in the centres of these modules. See Supplementary Figures S13-S16 for further co-expression results.
We used these co-expression modules for pathway analyses as described in Methods. The importance of extracellular matrix, vascular smooth muscle cell contraction and developmental pathways were highlighted by enrichment of GO biological pathway and molecular function terms (e.g. extracellular matrix organisation, collagen-containing extracellular matrix, contractile fibre, muscle contraction, actin binding, myosin binding, cardiovascular system development). GO terms related to the TGF-β pathway were also significantly enriched in functional modules derived from gene associations with all the aortic phenotypes (FDR<0.05).
Phenome-wide association studies and MR-PheWAS
We performed a Phenome Wide Association Study (PheWAS) and a subsequent Mendelian Randomisation – Phenome Wide Association Study (MR-PheWAS) to explore associations between our aortic traits of interest and clinical diagnoses in the whole UK Biobank population for which relevant clinical data were available (n= up to 406,827), controlling for age and sex.
Initial PheWAS identified significant phenotypic associations between aortic traits and multiple hypertension-related clinical codes. Aortic areas showed a positive phenotypic association with hypertension (for example, AAmin β=0.001; p=2.5×10−21). Aortic distensibility was negatively associated with hypertension (β=-0.217; p= 1.65×10−25). There was a significant negative association for all traits with type II diabetes mellitus.
A subsequent analysis of genotypic associations using MR-PheWAS supported a significant causal relationship between AAdis (MTAG) and aortic aneurysms (Inverse Variance Weighted (IVW) OR=0.28, 95% CI 0.16-0.50; p value 2.14×10−5; consistent directions of effect with Weighted Median (WM)/MR Egger and with use of the single trait analysis of AAdis as the genetic instrument, though the latter did not reach significance), suggesting clinical meaningfulness of the distensibility phenotype. MR-PheWAS also suggested that ascending and descending aortic areas are causally related to the risk of aortic aneurysms without evidence of significant pleiotropy.
Relationship between blood pressure and aortic dimensions
We tested further for bi-directional causal relationships between quantitative blood pressure traits (using GWAS summary statistics from a previous study35) and aortic areas using Mendelian randomisation (MR). MR results supported a bidirectional causal relationship between ascending aortic areas and diastolic blood pressure (DBP; AAmin->DBP; βIVW=0.004, p=4.3×10−16; DBP->AAmin: βIVW=6.6; p=2.5×10−5) and between ascending aortic area and pulse pressure (PP; AAmin->PP: βIVW=-0.007; p=7.1×10−15; PP-> AAmin: βIVW=-8.1, p=4.4×10−7). MR-Egger estimates were consistent for all but the DBP-> AAmin analysis, for which the estimates were in the opposite direction. Contamination mixture MR (MR-ConMix, see Methods) showed consistent findings for all analyses. Similar analyses for causal relationships with blood pressure were not performed for distensibility since blood pressure is used for calculation of the trait.
Genetic relationships between aortic traits and cerebral small vessel disease or cervical artery dissection
We explored genetic correlations and potential causal relationships between aortic traits and brain SVD estimated from the brain MRI measure of white matter hyperintensity (WMH) burden. Using LDSC, we identified a significant genetic overlap between all aortic traits and WMH burden which defined a positive association with minimum aortic area and an inverse association for the distensibility measures (AAmin rg=0.20, p=0.001; DAmin rg=0.22, p=2.19×10−5; AAdis rg=-0.22, p=5.0×10−4; DAdis rg=-0.33; p=1.0×10−4). In further analyses we found no significant genome-wide overlap between aortic traits and risks of cervical artery dissection, a leading cause of stroke in young people which was associated with aortic phenotypes in an earlier study36. However, the regional level overlap estimates from a Bayesian pairwise GWAS (GWAS-PW) suggest a high probability of shared variants between aortic distensibility traits and the cervical artery dissection (CeAD) risk locus PHACTR1-EDN1. Most of the other CeAD risk loci with high probability of shared variants with aortic traits harbour single nucleotide variants (SNVs) associated at genome-wide significance with one or more of the aortic traits. Exceptions to this include regions at chromosomes 12 (including c12orf49, RNFT2, PAWR, OTOGL), 16 (including CMIP, PKD1L2, BCO1) and 2 (including MBD5, EPC2, LYPD6B), suggesting further novel biologically relevant associations with aortic traits in these regions.
Exploring relationships between aortic traits and white matter hyperintensity burden using Mendelian randomisation
A lower aortic distensibility or a greater ascending aortic area is genetically correlated with increased burden of WMH in our data. We hypothesised that there might be a causal relationship between these aortic traits and cerebral small vessel disease. Although a two-sample MR using genetic associations with aortic traits as the instrumental variable and WMH as the outcome showed no evidence for a causal association for any of the aortic traits after FDR correction, after accounting for the effect of blood pressure (either systolic or pulse pressure) using a multivariable MR, we found evidence for a direct causal effect of both ascending and descending aortic distensibility and ascending aortic area on WMH burden. Lower distensibility and higher area were associated with an increased WMH burden (for AAdis β=-0.12, p= 1.49×10−3 and DAdis β=-0.21, p=1.14 ×10−3 using MTAG-derived associations as the instrumental variable and for AAmin β=3.8×10−4, p=2.91×10−3 using stage 1 associations as the instrumental variable).
DISCUSSION
Our analysis provides a first large-scale GWAS of ascending and descending aortic distensibilities, and adds substantively to the literature concerning the genetic basis of variation in aortic dimensions. We show that aortic distensibility has a significant heritable component, with 11% of the variance in AA distensibility and 10% of the variance in DA distensibility explained by the common genetic variants included in our study (increasing to 24% and 21% respectively for the MTAG analysis). We identify 38 loci associated with these measures of aortic stiffness, and a total of 31 novel loci for aortic traits including aortic areas (102 loci overall). Annotation of these loci provides evidence for mechanistic associations of TGF-β, IGF, PDGF and VEGF signalling pathways with aortic distensibility.
The clinical significance of our findings is suggested by the potential causal associations between AAdis (and other aortic traits) and aortic aneurysms defined by MR-PheWAS. Multivariable MR provides new evidence for possible mechanistic associations between cerebral small vessel disease and both aortic distensibilities and aortic area, helping to explain their long-recognised clinical relationships20, 37.
Mendelian aortopathy or cardiovascular disease associated genes ELN38, THSD439, FBLN540, 41, PRDM642 and ABCC943 directly overlap loci associated with aortic distensibility phenotypes and thus are strong candidates for expression of functional effects of variation at the corresponding loci. Similarly, Mendelian disease genes FBN144, MYH745, TBX2046, MASP147 and LOX48 overlap loci associated with aortic area phenotypes. Other genes associated with aortic area in our analyses have previously been associated with risks of acute aortic dissection (ULK4, LRP149). Several gene ontologies were associated with our measured aortic traits which are also of significance in Mendelian aortic disease50. Those GO terms related to the extracellular matrix, cardiovascular development and vascular smooth muscle cell function (with genes such as ELN, ABCC9, ANO1 and PRDM6 associated with AAdis) were amongst the most consistently identified. This overlapping genetic landscape of distensibility (and aortic areas) and aortic aneurysms, and our finding of a likely causal link between these phenotypes using MR-PheWAS suggest that functional pathways related to genes associated with quantitative aortic traits contribute to the pathogenesis of aortic disease. Together, these observations also support the growing consensus that cardiovascular disease phenotypes may be expressed as a result of extremes of normal genetic variation in the population59, and support the use of distensibility to predict both aneurysm formation and progression4.
Our data also support a role for TGF-β signalling in determining aortic distensibility in the general population. The TGF-β pathway has long been recognised as an important modulator of aortic function, with variants in many of the major components identified as causal in Mendelian aortic disease such as Loeys-Dietz syndrome51. Gene ontologies and cell-specific co-expression modules associated with the measured aortic traits suggest that IGF, PDGF and VEGF signalling also play significant roles in aortic biology. The roles of insulin and of insulin-like growth factor signalling are of considerable therapeutic interest for aortic pathology given recent evidence suggesting that metformin, a known regulator of both signalling pathways54, could be an effective treatment for abdominal aortic aneurysm, and the consequent initiation of clinical trials testing this55, 56.
Each of these gene sets offers interesting candidates in the search for new Mendelian aortic disease genes. Our results also add to the literature on sex differences in the genomic regulation of cardiovascular traits57, 58, with new evidence presented here suggesting distinct, biologically relevant associations in males and females and implicating genes such as ADAMTS7, SCAI, ALDH2 and FES as sex-specific determinants of aortic traits and thus possibly also the related diseases.
The strongest SNV association with ascending aortic distensibility was found in close proximity and upstream of ELN, the gene encoding elastin, a functionally central component of aortic elastic fibres. FBLN5 was strongly associated with DAdis. This encodes fibulin-5, another key extracellular matrix protein and a mediator of elastic fibre assembly63. Variants in FBLN5 cause a Mendelian form of cutis laxa associated with aortic aneurysm, vascular tortuosity and supravalvular aortic stenosis40, 41. The importance of extracellular matrix (ECM) composition and regulation is also demonstrated by the identification of multiple ECM-related GO terms associated with aortic phenotypes. Specific gene associations also serve to emphasise this, including three members of the ADAMTS family, which regulate ECM turnover: ADAMTS7 and ADAMTS8, both previously associated with aortic minimum areas and replicated here, and a novel association of ascending aortic distensibility with ADAMTS9. The strong association of AAdis with HAS2 (encoding a hyaluronan synthetase) demonstrated that glycosaminoglycan components of the ECM are also key determinants of aortic traits.
Other specific associations provide insights into the complexity of aortic biology. For example, the second most significant association with AAdis is within a long, non-coding RNA (lncRNA) just 3’ of PCSK1, a proprotein convertase whose substrates include many hormones such as renin, insulin and somatostatin (associated previously with body mass index and obesity64) and therefore which may mediate multiple endocrine influences on aortic traits. The third most significant locus associated with AAdis was previously associated with coronary artery disease65 and spans SMG6 (a regulator of nonsense mediated decay) and SRR, a serine racemase. The causal gene at this locus is thought to be SMG6, although functional data demonstrating strong eQTLs for SRR in all the risk alleles identified suggests it remains a candidate gene for this locus.
The shared genetic basis of ascending and descending distensibilities is limited (Supplementary Figure 4), consistent with the differing developmental origins of these parts of the aorta, and associated differences in structures of the aortic wall, in which elastin content diminishes more distally66. The most significant association for descending aortic distensibility is in PLCE1, a gene previously associated with blood pressure traits35, 67. Evidence from knockout mice suggests that PLCE1 contributes to the integration of β-adrenergic signalling with inputs from IGF-1 and other pathways to regulate cardiomyocyte differentiation and growth. We speculate that it might play a similar integrative role in the development and remodelling of aortic tissues.
Associations between aortic traits and brain small vessel disease have long been recognised, but the mechanisms responsible have not been defined clearly20, 68. This has been a particularly difficult relationship to untangle, as both are subject to confounding influences of blood pressure and other pleotropic factors. Our multivariate MR provides novel evidence suggesting that aortic traits including distensibility are causally linked to WMH and that this relationship is independent of (and additive to) the effects of blood pressure. By inference, as WMH burden predicts cognitive decline and dementia (with evidence supporting a causal association with Alzheimer type dementia16, 37), these results indirectly suggest that aortic stiffening could also contribute to cognitive decline and dementia, e.g. through altered haemodynamics and resultant changes in cerebral blood flow leading to effects on brain endothelial cell function and small vessel remodelling69, 70.
A shared genomic influence on aortic distensibility and cervical artery dissections was identified at the PHACTR1/EDN1 locus. Previously, this locus was implicated in coronary artery disease71, myocardial infarction72, migraine73, fibromuscular dysplasia74 and cervical artery dissection75. We demonstrated an association of EDN1 with ascending aortic distensibility, and further characterised EDN1 as a hub gene in co-expression networks derived from aortic endothelia, suggesting that EDN1 may be responsible for (or functionally contribute to) this shared genetic association with both CeAD risk and aortic distensibility,
Although we have made several novel observations, there are obvious limitations of our study. The GWAS was restricted to the analysis of Caucasian individuals, and so the generalisability of our results for people from other ethnic backgrounds is uncertain. Second, the accuracy (and possibly also the precision) of the distensibility measures likely was reduced by the need to use non-invasive blood pressure measurements (acquired on the same day as the imaging) as proxies for central blood pressure recordings. The ascending and descending aortic distensibility measures also suffer from confounding due to a likely bidirectional relationship with blood pressure, given the use of blood pressure indices in the derivation of the phenotype. Uncontrolled confounding from residual effects of blood pressure could bias the Mendelian randomization analyses. Nevertheless, our genetic associations were significantly enriched for genes expressed in the aorta and identify genes known to be important in aortic biology, affording some confidence in the robustness of our results.
In summary, our results provide genetic association data highlighting roles for TGF-β and other growth factor (IGF, PDGF, VEGF) signaling pathways in the elastic function of the aorta and, by inference, in aortic disease. We present new evidence for potential causal links between lower aortic distensibility and increased risk of aortic aneurysm and for common causal mechanisms relating cerebral small vessel disease and aortic structure and function that could explain the clinically observed relationships between late life cognitive decline and aortic disease14, 18. Better understanding of the underlying mechanisms based on these genetic data could lead to the identification of new therapeutic targets for reduction of both cardiovascular disease and dementia risks.
METHODS
Data
The UK Biobank cardiac magnetic resonance imaging (CMR) was conducted using a rigorously controlled acquisition protocol76. The mean age at the time of CMR was 64 ± 8 years (range 45–82, 49% of participants were male). Exclusion criteria for imaging included a range of relative contraindications to magnetic resonance imaging scanning as well as childhood onset disease and pregnancy. Aortic cine images were acquired using transverse bSSFP sequence at the level of the pulmonary trunk and right pulmonary artery on clinical wide bore 1.5 Tesla scanners (MAGNETOM Aera, Syngo Platform VD13A, Siemens Healthcare). Each cine image sequence consists of 100 time frames. The typical image size is 240×196 pixel with the spatial resolution of 1.6×1.6 mm2. Brachial blood pressure was obtained using a manual sphygmomanometer and converted into central blood pressure for the distensibility calculations by applying a brachial-to-aortic transfer function using the Vicorder software76.
Derivation of imaging phenotypes
A recurrent convolutional neural network was developed for aortic image segmentation and trained using manual annotations of 800 ascending aorta and descending aorta images (400 subjects and 2 time frames per subject)25, 27 The network was applied to segmenting aortic images across the cardiac cycle. A semi-automated quality control was performed for all segmentations, consisting of automated checking of missing or fragmented segmentation and subsequent manual checking on segmentation screenshots. Six aortic imaging phenotypes were calculated based on the automated segmentations, including those for the maximal area, minimal area and distensibility for both the ascending aorta and descending aorta4. Distensibility was calculated as
 where Amax and Amin denote the maximal and minimal area and SBP and DBP denote the systolic and diastolic central blood pressure, measured at the imaging visit during the study protocol (measured brachially and converted to central blood pressure by applying a brachial-to-central transfer function as described above). After running the image segmentation pipeline and performing quality control, imaging phenotypes were available for 36,996 participants.
where Amax and Amin denote the maximal and minimal area and SBP and DBP denote the systolic and diastolic central blood pressure, measured at the imaging visit during the study protocol (measured brachially and converted to central blood pressure by applying a brachial-to-central transfer function as described above). After running the image segmentation pipeline and performing quality control, imaging phenotypes were available for 36,996 participants.
Genomic analyses
We performed stage 1 GWAS on six imaging phenotypes (AAmax, AAmin, DAmax, DAmin, AAdis and DAdis). Outliers with phenotype values >4 standard deviations (SDs) from the mean were excluded to ensure we did not include patients with undiagnosed aortic aneurysm in our results. After exclusions for image quality control, outlying BMI (<15 or >40), stage IV hypertension, missing covariates, diagnosis of aortic disease and non-white ethnicity, 4,406 participants were excluded leaving 32,590 individuals for the GWAS of AAmax, AAmin, DAmax, and DAmin, and 29,895 for the GWAS of AAdis and DAdis (the latter figure is lower due to more missing contemporaneous blood pressure recording data and incomplete imaging sets). See Supplementary Figure S1 for more details on exclusions. After exclusions, we rank-normalised the distensibility phenotypes due to the non-normal distribution of the distensibility phenotypes (Supplementary Figure S2). Aortic area phenotypes approximated a normal distribution and so raw areas were used to facilitate interpretation of the effect sizes. The genetic model was adjusted for age at time of imaging, sex, mean arterial pressure, height, and weight. We used the linear mixed model approach implemented in BOLT-LMM (v2.3.4)77. The genetic relationship matrix (GRM) constructed by BOLT was based on all directly genotyped SNPs (N = 340,336) passing the threshold settings (MAF > 0.05, p(HWE) > 1e-6 and genotype calling rate > 98.5). For the main analysis, a threshold of MAF >0.01 was applied to the SNPs. Genomic inflation (lambda) was calculated in R as the median chi square values derived from the p-values divided by the expected median of a chi square distribution with 1 degree of freedom.
We used MTAG (version 1.0.8)28 for multi-trait analysis of GWAS summary statistics to increase power. MTAG can identify genetic loci associated with a particular phenotype where the single-trait GWAS is underpowered. The method uses the correlation structure of the trait in question with other traits to boost power. It therefore may fail to identify trait-specific loci but will increase power to detect loci associated with the other related and correlated traits. We used all 6 aortic phenotypes for our MTAG analysis. Regression coefficients (beta) and their standard errors were used for MTAG. The results of the multi-trait analyses are shown as Figure 1B and in Supplementary Figures S7-8.
We additionally conducted a sex-specific analysis by performing GWAS on aortic areas for autosomal SNPs in men and women separately, using the BOLT-LMM pipeline as above, and compared the sizes of the sex-specific genetic associations for SNPs with a P-value smaller than 5×10−8 using a z-test78. We did not repeat this analysis for distensibility phenotypes as it was underpowered because of the reduced cohort size and smaller effect sizes for the distensibility SNPs as well as lower heritability estimates for these traits.
To classify genomic loci associated with our imaging phenotypes, GWAS summary statistics were processed using FUMA (v1.3.6)32 and a pre-calculated LD structure based on the European population of the 1000 Genome Project82. SNPs that reached genome-wide significance (p<5×10−8) and with r2<0.6 were defined as independently significant. All variants with r2 ≥ 0.6 were labelled as candidate variants for further annotation by FUMA. In a second clumping procedure to define lead SNPS, those correlated with r2<0.1 were defined as independently significant. Finally, proximal LD blocks of independent significant SNPs with less than 250kb distance were merged and considered as a single genomic locus. To consolidate genomic loci across different traits, the merge function implemented in bedtools was applied79.
For the association of SNPs with genes, biological processes and tissue expression, we applied the SNP2GENE function implemented in FUMA to the summary SNP statistics. For the positional mapping of SNPs to genes, a maximal distance of 10kB was set. eQTL mapping was performed based on the aorta tissue samples in GTEx v8 using only gene pairs with significant SNPs (with the default settings of FDR < 0.05 or p value < 1e-3). 3D chromatin interaction mapping was based on HiC aorta data (GSE87112) within a promotor region window of 250bp upstream and 500bp downstream from the transcriptional start site and a threshold for significant loops of FDR < 1e-6. Enhancer and promotor regions were annotated using the aorta epigenome (E065) from the Roadmap Epigenome Project (http://www.roadmapepigenomics.org/). MAGMA (Multi-marker Analysis of GenoMic Annotation) was employed to obtain the significance of individual genes with the specificity of tissue expression based on 54 types in GTEx v8 and the association with 10,678 gene sets from MsigDB v6.2 (with 4761 curated gene sets and 5917 GO categories). Further annotation of significant SNPs and loci was performed manually with SNP lookups in GTEx v829; normalised effect sizes are reported from this dataset using “Artery-Aorta” for the main analysis and including other arterial tissue types (“Artery – Tibial” and “Artery – Coronary”) where stated. For Gene Ontology (GO) enrichment analysis, the Data-driven Expression Prioritized Integration for Complex Traits (DEPICT) software was applied (v1 beta rel194, www.broadinstitute.org/depict), which is based on probabilistic memberships of genes across reconstituted gene sets80 For LD-based clumping by PLINK81, which precedes the DEPICT analysis, a p-value threshold of 10−5, a distance threshold of 500kb and a LD threshold of 0.1 was set (following the recommendations on the DEPICT website). Note that DEPICT excludes any SNPs in the HLA region, on a sex chromosome, or not found in the 1000 Genomes Project data.
Comparisons between our data and those reported by Pirruccello et al30. and Tcheandjieu et al31 were made using the lead SNPs and corresponding beta-values. To assess the degree of convergence of their studies with our results, lead SNPs were assigned to a genomic locus found in our study if they were within the locus or a 250kb distance interval.
LD Score regression
We performed LD Score regression to assess the heritability of the imaging phenotypes. The genetic correlation between imaging phenotypes and blood pressure by LD score was computed using 1000 Genomes European data82. We used the GWAS summary statistics for SBP, DBP and pulse pressure (PP) from the International Consortium for Blood Pressure (ICBP)35 for the corresponding analyses for genetic correlation with blood pressure.
Co-expression analysis
Co-expression networks for aortic phenotypes were derived using a single cell RNA-seq (scRNA-seq) data set for primate arteries34. The data were obtained from Gene Expression Omnibus (accession number GSE117715) and included read counts for over 9000 single cells from aortas and coronary arteries of 16 Macaca fascicularis. Low abundance genes were removed if they had read counts of less than 5% of the cells, leaving a total of 9903 genes for further analysis. The Bioconductor package scater was applied to compute log-transformed normalised expression values from the count matrix83. Subsequently, correlation of expression and its significance was derived using the correlatePairs function of the Bioconductor scran package, which calculated modified Spearman correlation coefficients and derived their significance using a permutation approach84. A basal co-expression network was constructed using gene pairs with significant correlation (FDR < 0.01) and a minimum Spearman correlation coefficient rho of 0.1 or 0.2. Subsequently, sub-networks for imaging phenotypes were derived by retrieving gene pairs with at least one gene associated by MAGMA or FUMA with the specific phenotype.
Functional enrichment of genes connected with variants identified on GWAS was carried out using overrepresentation enrichment analysis implemented in the Bioconductor clusterProfiler package85. The background gene set (or universe) was defined by the genes covered by the scRNA-seq data after exclusion of low abundance genes.
Mendelian randomization (MR)
To investigate potentially causal relationships between aortic phenotypes and diseases, we performed bidirectional two-sample Mendelian randomization (MR) using our GWAS for aortic imaging phenotypes and the International Consortium for Blood Pressure GWAS on blood pressure35. We limited our MR analyses to AAmax, AAmin, DAmax, and DAmin because blood pressure is included in the calculation of the distensibility measure.
For either direction of potential causal relationships between the aortic phenotypes and blood pressure, we selected SNPs associated with the exposure at genome-wide significance level (P-value<5×10−8 and F-statistic>10). For SNPs correlated with r2 greater than 0.1, we used only the SNP with the smallest p-value for the SNP-exposure association. We tested the validity of the genetic variants as instrumental variables using the contamination mixture method (MR-ConMix)86. The contamination mixture method constructs a likelihood function based on the SNP-specific estimates and evaluates the SNP-specific contribution to the likelihood.
In each case, we estimated SNP-specific associations as the ratio of SNP outcome to exposure associations (Wald ratio)87. SNP-specific associations were combined using the inverse-variance weighted (IVW) estimator 88. The putative causal effect (βIVW) of an exposure on a given outcome was estimated using the inverse-variance weighting (IVW) method as the weighted sum of the ratios of beta-coefficients from the SNP-outcome associations for each variant (j) over corresponding beta-coefficients from the SNP-exposure associations (βj). The ratio estimates from each genetic variant were averaged after weighting on the inverse variance (Wj) of βj across L uncorrelated SNPs,
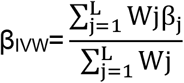
We also used weighted median (WM) and MR-Egger regressions as sensitivity methods to test the robustness of associations 88. Potential horizontal pleiotropic effects were investigated using MR-Egger 89. Outlier SNPs identified by MR-PRESSO were excluded from the analyses 90. In an additional analysis, we tested the validity of the genetic variants as instrumental variables using MR-ConMix86. We accounted for multiple comparisons of four aortic imaging phenotypes, three blood pressure traits, and two directions using Bonferroni correction with a P-value threshold of 0.05/(4*3*2)=0.002
MR analysis was also used to investigate the potential causal relationships between different aortic traits with WMH. In addition to IVW, WM, and MR-Egger, we implemented R package RadialMR (available through CRAN repositories)91. A p-value < 0.01 correcting for 6 tests (for the 6 aortic traits) was considered significant. In the presence of heterogeneity (PHet<0.01, Cochran’s Q statistic) due to horizontal pleiotropy, RadialMR was used in the identification of pleiotropic SNPs that have the largest contribution to the global Cochran’s Q statistic by regressing the predicted causal estimate against the inverse variance weights. After excluding influential outlier SNPs, the IVW test was repeated along with MR-Egger regression in which the regression model contains the intercept term representing any residual pleiotropic effect 92. Non-significant MR-Egger intercept was used as an indicator to formally rule out horizontal pleiotropy. Relative goodness of fit of the MR-Egger effect estimates over the IVW approach was quantified using QR statistics, which is the ratio of the statistical heterogeneity around the MR-Egger fitted slope divided by the statistical heterogeneity around the IVW slope. A QR close to 1 indicates that MR-Egger is not a better fit to the data and therefore offers no benefit over IVW 91.
Multivariable MR
We also conducted multivariable two sample MR (MVMR) to estimate the direct effect of aortic traits on the cerebral small vessel disease (cSVD) outcome (WMH) after accounting for potential confounding with blood pressure traits, by conditioning on every other explanatory variable included in the model. Different combinations of explanatory variables were considered and the FTS conditional on the other variables was calculated as a measure of instrument strength 93. Briefly, MVMR by regressing a given instrumental variable on all the remaining variables as controls generates a predicted value for the instrumental variable that is not correlated with other variables in the model thus accounting for possible pleiotropic effects.
MR phenome-wide association studies (MR-PheWAS)
We performed an MR-PheWAS using data from UK Biobank participants who did not undergo aortic imaging to assess the effects of aortic traits on clinical disease classifications. Using the PheWAS package (https://github.com/PheWAS/PheWAS), we mapped 1,157 phecodes with more than 200 cases from the International Classification of Diseases, 9th Revision, Clinical Modification (ICD-9-CM) and the International Classification of Diseases, 10th Revision, Clinical Modification (ICD-10-CM).94 For each aortic imaging phenotype, we selected SNPs with a p-value<5×10−8 and minor allele frequency > 0.05 from the single trait GWAS for AAmax, AAmin, DAmax, and DAmin, and from the multi trait GWAS for AAdis and DAdis. Given that we considered a large number of phecodes as the outcome in the MR-PheWAS analysis with the number of cases ranging from 201 to 116,879 (median=1,353), we used a more stringent LD threshold for independent SNPs (r2<0.01) to achieve a better stablised model. The PheWAS model was adjusted for age, sex, genotype array, and its four principal components for population stratification. MR estimates were then obtained for each pair of aortic traits and phecode by combining the SNP-specific associations using IVW, WM, and MR-Egger. We accounted for multiple comparisons of 1,157 phecodes using Bonferroni correction with p-value<0.05/1,157 =4.3×10−5.
Analyses of associations of aortic and cervical artery dissection
LD score regression (LDSR) method was applied to test genetic correlation at the genome-wide scale for the different aortic traits with the most common MRI feature of cSVD, WMH and with cervical artery dissection (CeAD). GWAS summary statistics were obtained from recently published consortia GWAS of cerebral phenotypes (from the CHARGE and CADISP consortia respectively)37,75. For this, common variants, mapping to the Hapmap3 reference panel were employed. As the slope from the regression of the Z-score product from the two GWAS summary statistics on the LD-score gives the genetic covariance, the intercept of the genetic covariance was used as an indirect measure of sample overlap 95, which corresponds to the average polygenic effects captured by genetic variants spread across the genome. A p-value < 0.006 (adjusting for 8 simultaneous tests) was considered significant.
LDSR could potentially miss significant correlations at the regional level due to the balancing-effect96. A Bayesian pairwise GWAS approach (GWAS-PW) was applied to systematically test for locally correlated regions97. The GWAS-PW identified trait pairs with high posterior probability of association (PPA) using a shared genetic variant (model 3, PPA3> 0.90). To ensure that PPA3 is unbiased by sample overlap, fgwas v.0.3.698 was run on each pair of traits and the correlation estimated from regions with null association evidence (PPA3<0.20) was used as a correction factor. Additionally, to estimate the directionality of associations between trait pairs in regions with PPA3>0.90, a simple rank-based correlation test was applied. Independence between regions was estimated as proposed by Berisa and Pickrell99. Only the most strongly associated variant for the outcome per region showing high PPA3 is reported.
Data Availability
Summary statistics for all GWAS analyses are available upon request.
SUPPLEMENTARY FIGURES
Supplementary Figure 1 Exclusions
Supplementary Figure 2 Distribution of traits
Supplementary Figure 3 Phenotypic correlation between traits
Supplementary Figure 4 Genotypic correlation between traits (LDSC)
Supplementary Figure 5 All Manhattan plots stage 1 GWAS
Supplementary Figure 6 All QQ plots stage 1 GWAS
Supplementary Figure 7 All Manhattan plots MTAG
Supplementary Figure 8 All QQ plots stage 2 (MTAG) GWAS
Supplementary Figure 9 FUMA genes results
Supplementary Figures 10 MAGMA genes Manhattan plots
Supplementary Figure 11 MAGMA tissue expression
Supplementary Figure 12 MAGMA GO heatmap
Supplementary Figure 13 Coexpression networks for AAdis
Supplementary Figure 14 Coexpression networks for DAdis
Supplementary Figure 15 Coexpression networks for AAmin
Supplementary Figure 16 Coexpression networks for DAmin
CADISP list of investigators
Belgium: Departments of Neurology, Erasmus University Hospital, Brussels and Laboratory of Experimental Neurology, ULB, Brussels (Shérine Abboud, Massimo Pandolfo); Department of Neurology, Leuven University Hospial (Vincent Thijs). France: Departments of Neurology, Lille University Hospital-Inserm U1171 (Didier Leys, Marie Bodenant), Sainte-Anne University Hospital, Paris (Fabien Louillet, Emmanuel Touzé, Jean-Louis Mas), Pitié-Salpêtrière University Hospital, Paris (Yves Samson, Sara Leder, Anne Léger, Sandrine Deltour, Sophie Crozier, Isabelle Méresse), Amiens University Hospital (Sandrine Canaple, Olivier Godefroy), Dijon University Hospital (Maurice Giroud, Yannick Béjot), Besançon University Hospital (Pierre Decavel, Elizabeth Medeiros, Paola Montiel, Thierry Moulin, Fabrice Vuillier); Inserm U744, Pasteur Institute, Lille (Jean Dallongeville). Finland : Department of Neurology, Helsinki University Central Hospital, Helsinki (Antti J Metso, Tiina Metso, Turgut Tatlisumak); Germany: Departments of Neurology, Heidelberg University Hospital (Caspar Grond-Ginsbach, Christoph Lichy, Manja Kloss, Inge Werner, Marie-Luise Arnold), University Hospital of Ludwigshafen (Michael Dos Santos, Armin Grau); University Hospital of München (Martin Dichgans); Department of Rehabilitation: Schmieder-Klinik, Heidelberg (Constanze Thomas-Feles, Ralf Weber, Tobias Brandt). Italy: Departments of Neurology: Brescia University Hospital (Alessandro Pezzini, Valeria De Giuli, Filomena Caria, Loris Poli, Alessandro Padovani), Milan University Hospital (Anna Bersano, Silvia Lanfranconi), University of Milano Bicocca, San Gerardo Hospital, Monza, Italy (Simone Beretta, Carlo Ferrarese), Milan Scientific Institute San Raffaele University Hospital (Giacomo Giacolone); Department of Rehabilitation, Santa Lucia Hospital, Rome (Stefano Paolucci). Switzerland: Department of Neurology, Basel University Hospital (Philippe Lyrer, Stefan Engelter, Felix Fluri, Florian Hatz, Dominique Gisler, Leo Bonati, Henrik Gensicke, Margareth Amort). UK: Clinical Neuroscience, St George’s University of London (Hugh Markus). USA : Department of Neurology, Salt Lake City, USA (Jennifer Majersik); Department of Neurology, University of Virginia, Charlottesville, USA (Bradford Worrall, Andrew Southerland); Department of Neurology, Baltimore, USA (John Cole, Steven Kittner)
COMPETING INTERESTS/DISCLOSURES
PMM acknowledges consultancy fees from Novartis, Bristol Myers Squibb, Celgene and Biogen. He has received honoraria or speakers’ honoraria from Novartis, Biogen and Roche and has received research or educational funds from Biogen, Novartis, GlaxoSmithKline and Nodthera.
CO-AUTHOR CONTRIBUTIONS
CF, MF, JH, SF, AD and PMM co-designed the study. CF, MF, WB, JH, AP, AR and MS performed quality control and core analyses. SD, EP, JW and PE provided additional methodological guidance. CF, MF, JH, AD and PMM drafted the manuscript. All authors reviewed, contributed to serial revisions and approved the manuscript.
ACKNOWLEDGEMENTS
PMM acknowledges generous personal and research support from the Edmond J Safra Foundation and Lily Safra, a National Institute for Health Research (NIHR) Senior Investigator Award, the UK Dementia Research Institute and the NIHR Biomedical Research Centre at Imperial College London. CF acknowledges generous support from the BHF Imperial Centre of Research Excellence RE/18/4/34215. This research has been conducted using the UK Biobank Resource under Application Number 18545. PE is director of the Medical Research Council (MRC) Centre for Environment and Health and acknowledges support from the MRC (MR/L01341X/1; MR/ S019669/1). PE is director of the Health Protection Research Unit in Chemical and Radiation Threats and Hazards, funded by the NIHR. PE also acknowledges support from the NIHR Imperial Biomedical Research Centre, the Imperial College British Heart Foundation Centre for Research Excellence (RE/18/4/34215), the UK Dementia Research Institute at Imperial College London (MC_PC_17114), and Health Data Research UK for London. SD is supported by a grant overseen by the French National Research Agency (ANR) as part of the ‘Investment for the Future’ Programme ANR-18-RHUS-002 and by the ERC and the EU H2020 under grant agreements 640643, 667375, and 754517.
The CADISP study has been supported by Inserm, Lille 2 University, Institut Pasteur de Lille and Lille University Hospital and received funding from the ERDF (FEDER funds) and Région Nord-Pas de Calais in the frame of Contrat de Projets Etat-Region 2007-2013 Région Nord-Pas-de-Calais - Grant N°09120030, Centre National de Genotypage, Emil Aaltonen Foundation, Paavo Ilmari Ahvenainen Foundation, Helsinki University Central Hospital Research Fund, Helsinki University Medical Foundation, Päivikki and Sakari Sohlberg Foundation, Aarne Koskelo Foundation, Maire Taponen Foundation, Aarne and Aili Turunen Foundation, Lilly Foundation, Alfred Kordelin Foundation, Finnish Medical Foundation, Orion Farmos Research Foundation, Maud Kuistila Foundation, the Finnish Brain Foundation, Biomedicum Helsinki Foundation, Projet Hospitalier de Recherche Clinique Régional, Fondation de France, Génopôle de Lille, Adrinord, Basel Stroke-Funds, Käthe-Zingg-Schwichtenberg-Fonds of the Swiss Academy of Medical Sciences, Swiss Heart Foundation.
Footnotes
↵* joint first authors
References
- 1.↵
- 2.↵
- 3.
- 4.↵
- 5.↵
- 6.
- 7.
- 8.
- 9.
- 10.↵
- 11.↵
- 12.
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.
- 53.
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.
- 61.
- 62.
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
- 94.
- 95.↵
- 96.↵
- 97.↵
- 98.↵
- 99.↵



