
Abstract
Mortality due to Covid-19 is highly associated with advanced age, owing in large part to severe lower respiratory tract infection. SARS-CoV-2 utilizes the host ACE2 receptor for infection. Whether ACE2 abundance in the lung contributes to age-associated vulnerability is currently unknown. We set out to characterize the RNA and protein expression profiles of ACE2 in aging human lung in the context of phenotypic parameters likely to affect lung physiology. Examining publicly available RNA sequencing data, we discovered that mechanical ventilation is a critical variable affecting lung ACE2 levels. Therefore, we investigated ACE2 protein abundance in patients either requiring mechanical ventilation or spontaneously breathing. ACE2 distribution and expression were determined in archival lung samples by immunohistochemistry (IHC). Tissues were selected from the specimen inventory at a large teaching hospital collected between 2010-2020. Twelve samples were chosen from patients receiving mechanical ventilation for acute hypoxic respiratory failure (AHRF). Twenty samples were selected from patients not requiring ventilation. We compared samples across age, ranging from 40-83 years old in the ventilated cohort and 14-80 years old in the non-ventilated cohort. Within the alveolated parenchyma, ACE2 expression is predominantly observed in type II pneumocytes (or alveolar type II / AT2 cells) and alveolar macrophages. All 12 samples from our ventilated cohort showed histologic features of diffuse alveolar damage including reactive, proliferating AT2 cells. In these cases, ACE2 was strongly upregulated with age when normalized to lung area (p = 0.004) or cellularity (p = 0.003), associated with prominent expression in AT2 cells. In non-ventilated individuals, AT2 cell reactive changes were not observed and ACE2 expression did not change with age when normalized to lung area (p = 0.231) or cellularity (p = 0.349). Additionally, we observed prominent pulmonary endothelial ACE2 expression in 2 patients on either an angiotensin-converting enzyme inhibitor (ACEI) or angiotensin receptor blocker (ARB). In summary, ACE2 expression increases with age in the setting of alveolar damage observed in patients on mechanical ventilation, providing a potential mechanism for higher Covid-19 mortality in the elderly.
Introduction
The emergence of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has led to a global pandemic of related illness grouped under the term coronavirus disease 2019 (Covid-19)1. Manifestations include both upper and lower respiratory infection as well as various non-respiratory organ system disturbances2–6, including renal7, gastrointestinal8 and vascular abnormalities9. The vast majority of deaths result from lower respiratory tract infection leading to acute respiratory distress syndrome (ARDS) with the pathological findings of diffuse alveolar damage (DAD)10,11.
Risk factors for severe respiratory illness include hypertension12, diabetes12, obesity13, cancer14, chronic respiratory illness15, male sex13, and particularly advanced age16–18. For each year of age, the odds ratio of death has been estimated to increase by 1.1 in multivariable analysis16. Among individuals admitted to the ICU for Covid-19, risk of death for those > 64 years old is 2.4-fold greater than those younger19. In contrast, mortality among children infected with SARS-CoV-2 is exceedingly rare compared to middle-aged individuals20, a feature which is atypical for respiratory viruses21. The underlying reasons for this dramatic skew are currently unknown22.
SARS-CoV-2 depends on the angiotensin-converting enzyme 2 (ACE2) protein for attachment to and infection of host cells23. ACE2 is a transmembrane metalloprotease belonging to the Renin-Angiotensin-Aldosterone System (RAAS) which processes angiotensin II into angiotensin 1-7 leading to vasodilation24. The gene encoding ACE2 (Angiotensin I converting enzyme 2, Ace2) exhibits strong RNA expression in the kidney, lung, bladder, stomach, small intestine, large intestine, and adipose tissue of rodents25. In rats, lung ACE2 protein expression has been shown to drastically decrease with age26, raising the question of why older individuals are more susceptible to severe lower respiratory infection than the young27. Previous studies of ACE2 levels in the human lung have revealed no significant change with respect to age in bulk RNA sequencing (RNAseq) analysis28,29. Similar findings have been observed in single cell RNA (scRNA) expression data for Ace2 in rodents29. In order to investigate this apparent paradox, we reexamined publicly available ACE2 RNAseq data from human lungs30, adjusting for various donor phenotypes. Our analyses reveal a significant increase in detected ACE2 RNA levels associated with mechanical ventilation. After including the effect of mechanical ventilation, we find that ACE2 expression increases with age in human lung samples from deceased donors. Although RNA levels often correlate with protein expression, exceptions are common31,32. We sought to validate our findings at the RNA level by investigating ACE2 protein expression in archival human lung tissue from patients receiving supportive mechanical ventilation. Consistently, ACE2 protein expression strongly increased with age in patients requiring mechanical ventilation. These findings, at both the RNA and protein level, provide a potential explanation for the preponderance of severe Covid-19 cases among the elderly.
Methods
RNAseq analysis
Normalized gene expression data and subject phenotypes from the NIH Genotype-Tissue Expression (GTEx) project version 8 (released 8/26/2019) were acquired from the GTEx Portal33. These data include 17,382 samples from 948 deceased human donors. Expression scores in Transcripts Per Kilobase Million (TPM) were extracted for ACE2 from each sample and analyzed using R version 3.5.0. The expression for lung was plotted using ggplot2 stratified by donor age, sex, and/or the Hardy scale score (see Suppl. Methods), which are provided through the portal33. A linear model was fit using the lm function in base R to predict ACE2 expression with age, sex, and Hardy score as covariates. Primary analyses were carried out on linear TPM data to facilitate interpretation. The significance and direction of the effects were confirmed after Box-Cox transformation of lung ACE2 expression using λMLE computed by the MASS package in R. Mouse scRNAseq data34 for Ace2 expression were obtained from the Tabula Muris portal35 and droplet sequencing results for lung were plotted using ggplot2 in R.
Immunohistochemistry (IHC)
Formalin-fixed paraffin-embedded (FFPE) human lung excisions/biopsies received through Stanford Surgical Pathology during 2010-2020 were selected in accordance with an approved IRB protocol. To allow for blinding, cases were chosen by 1 investigator who did not participate in specimen preparation or data acquisition and analyses until all data had been collected. We identified a set of 20 patients who were not on mechanical ventilation (other than for procedural anesthesia) and a set of 11 patients who required supportive ventilation for acute hypoxic respiratory failure (AHRF); either idiopathic or in the setting of advanced fibrotic interstitial lung disease. Patients in our non-ventilated cohort ranged from 14 to 80 years of age (mean 41.6 +/- 23.1 years). Patients in our ventilated cohort ranged from 40 to 83 years old (mean 56.8 +/- 12.3 years). Sections were stained using standard procedures (see Suppl. Methods) with a commercial antibody raised against human ACE2 (Abcam, ab15348). Subsequent control sections were stained with Hematoxylin and Eosin (H&E) using standard procedures (Suppl. Figure 1A-D). All comparisons were made using sections that were stained in a single batch and imaged and processed using identical conditions (Suppl. Figure 2).
Image Collection and Quantification
Sections were imaged on a Nikon Eclipse E1000 using a Spot Insight 12 MP CMOS camera with the manufacturer’s provided software. A single inflated field within each slide was selected using a 1x objective from which to start image collection by switching to the 20x objective. Using the 20x objective, 5 adjacent/sequential fields were collected. Images used for quantification were obtained by an investigator without knowledge of the patients’ ages. ACE2 and cellular quantification were carried out using ImageJ (see Suppl. Methods). The relationship of age to these measures was analyzed by simple linear regression using the lm function in R.
Additional Statistical Analyses
Pairwise comparisons were made using the Wilcoxon rank-sum test. A threshold of p < 0.05 was set for statistical significance.
Results
To understand the age-related pattern of ACE2 abundance in the human lung we utilized the large, harmonized RNAseq dataset available from the GTEx consortium30. GTEx version 8 contains data from 578 unique deceased donor lungs, binned by decade of age at death33. In unadjusted analysis, ACE2 expression was not significantly associated with age (β = 0.082 TPM/decade, p = 0.227, linear regression) or sex (p = 0.303, Wilcoxon rank-sum test) (Figure 1A and 1B). The dataset also describes the mode of death, summarized by the Hardy scale36, which categorizes clinical circumstances proximate to the end of life. The strongest category associated with ACE2 expression was ventilation at the time of death showing a 103% increase in median ACE2 levels (0.715 TPM without vs. 1.452 TPM with ventilation, p < 1×10−15, Wilcoxon rank-sum test). Lung ACE2 expression, when stratified by Hardy score, revealed a significant relationship with age; expression of ACE2 increased by 0.208 TPM for every decade of life (p = 0.003, multivariable regression), or a 14% increase each decade when compared to individuals in their 20’s (Figure 1C). Transcript abundance from deceased donors can be affected by the tissue ischemic time that transpires prior to sample stabilization37. In order to assess whether this variable contributes to ACE2 abundance from deceased donor lungs, we incorporated this feature as a covariate in our model (Suppl. Figure 3). The contribution of tissue ischemic time to ACE2 expression was not significant (β = -0.0003 TPM/minute, p = 0.248) and did not change the effect of age on ACE2 expression or its significance (β = 0.212 TPM/decade, p = 0.002). We also compared ACE2 expression across the human body. Interestingly, when controlling for the manner of death, the lung exhibited the most significant increase in ACE2 expression with age over all tissues (Suppl. Table 1).
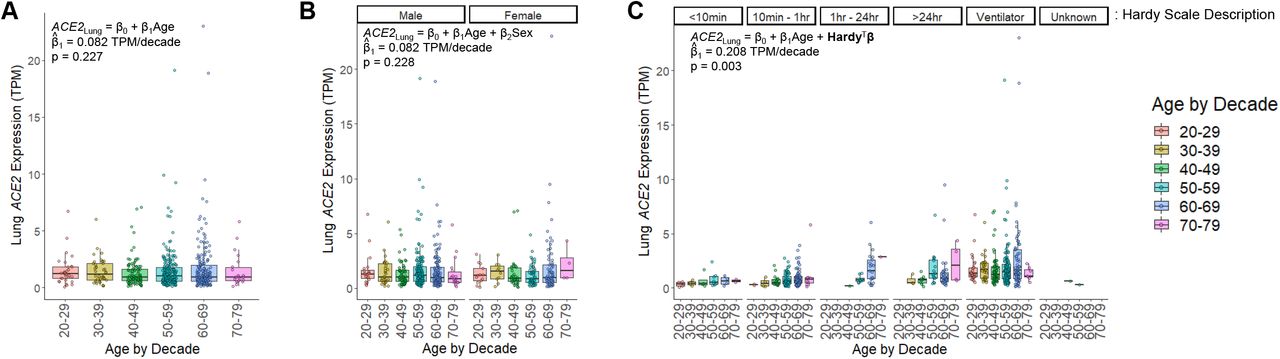
A) Human lung ACE2 expression is colored by decade of life when the individual died. In unadjusted linear regression, there was no significant effect of age on ACE2 expression (p = 0.227, n = 578 individuals). B) ACE2 expression in lung stratified by sex. Analysis adjusted for age and sex, revealed no significant effect for either variable (p = 0.228 for age, p = 0.920 for sex, multivariate regression, n = 395 males, n = 183 females). There was also no significant difference when compared by sex alone (p = 0.303, Wilcoxon rank-sum test). C) ACE2 expression in lung stratified by the Hardy scale. This scale indicates the length of time spent in the terminal phase before death, which is depicted above each grouping of data points. Within each Hardy scale group, the data are sub-stratified by age. (n = 26 for a score of 1 representing a violent and fast death lasting <10 minutes, n = 156 for a score of 2 representing a fast death by natural causes lasting 10 minutes – 1 hour, n = 31 for a score of 3 representing an intermediate rate of death lasting 1 hour – 24 hours, n = 64 for a score of 4 representing a slow death with a terminal phase lasting > 24 hours, n = 299 for a score of 0 representing donors supported by a ventilator preceding death, n = 2 with an unknown score). For all panels, each point represents a sample from a unique individual. Box plots indicate quartiles. A linear model fit to the data is inset, indicating the estimated coefficient for age (β1) and its significance.
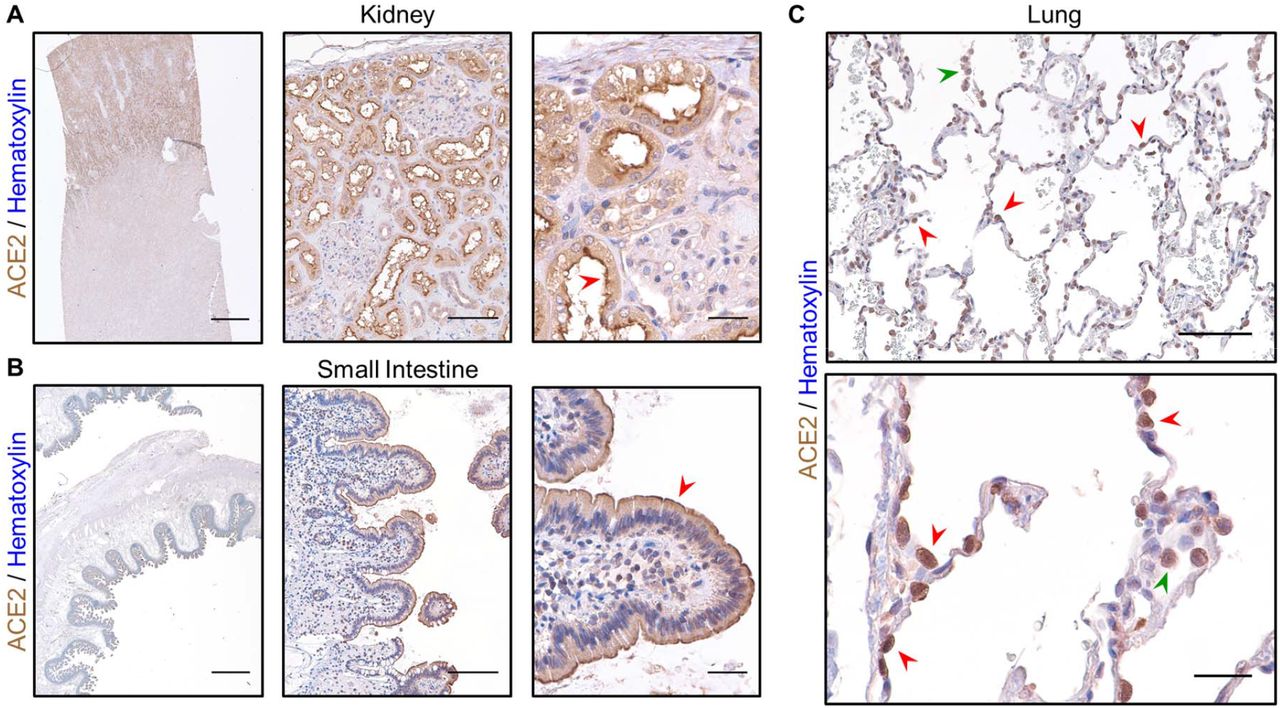
In order to characterize ACE2 protein abundance in the lung, we validated a commercial antibody directed against this key viral receptor (Figure 2A-C). Prior analysis of ACE2 localization by IHC revealed strong enrichment within the luminal surface of cortical tubule cells of the kidney and enterocytes of the small intestine38. Our ACE2 IHC staining on healthy control tissue revealed a strikingly similar pattern, which highlighted the kidney cortex and specifically the apical membrane of brush border cells (Figure 2A). We also detected strong ACE2 expression along villi and the apical membranes of small intestine enterocytes (Figure 2B). Within the lung, ACE2 has been shown to localize to epithelial cells of the alveolus38. Data from murine scRNAseq reveal detectable expression of Ace2 within type II pneumocytes (or alveolar type II / AT2 cells), ciliated columnar cells, alveolar macrophages, and to a lesser extent, stromal cells (Suppl. Figure 4). Consistent with these data, our IHC revealed strong ACE2 staining within AT2 cells in normal alveoli, concentrated within the membrane, as well as alveolar macrophages (Figure 2C). Taken together, these data indicate that our staining protocol accurately identifies ACE2 expressing cells within human tissue, including the lung. Therefore, we investigated whether the distribution of ACE2 in the lung is associated with ventilation and age.
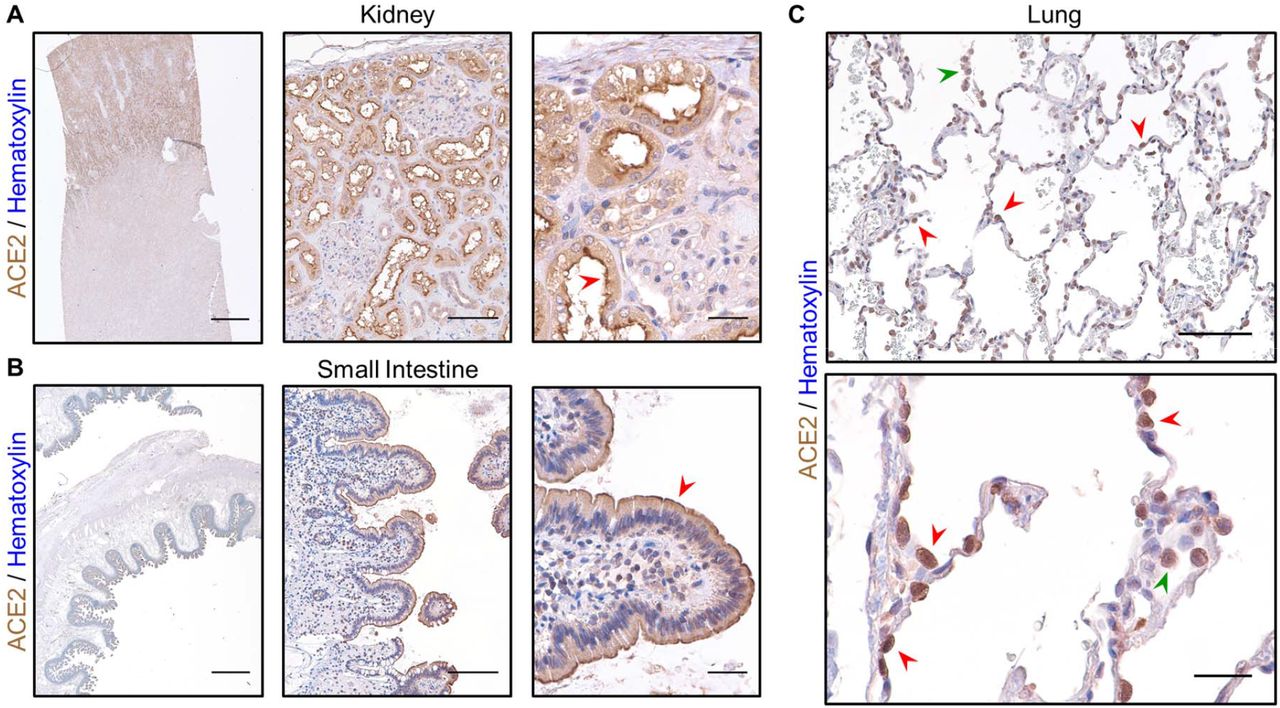
A) IHC staining for ACE2 in normal human kidney reveals strong signal in the cortex using a 1x objective (left, scale bar 3mm), within cortical tubules at low power (middle, scale bar 200µm), and within brush border cells at high power (right, scale bar 50µm) especially along the apical membrane (red arrowhead). B) ACE2 staining in normal human small intestine reveals signal within the luminal surface using a 1x objective (left, scale bar 3mm), along the epithelial lining of villi at low power (middle, scale bar 200µm), and in absorptive enterocytes at high power (right, scale bar 50µm) enriched in the apical membrane (red arrowhead). C) In normal human lung from a 23-year-old female, strong ACE2 expression is observed in AT2 cells (red arrowheads) along the alveolar septum at low power (top, scale bar 200µm) and in alveolar macrophages (green arrowhead). At high power (bottom, scale bar 50µm), ACE2 staining can be seen concentrated along the membrane of AT2 cells (red arrowheads) and within the cytoplasm of an alveolar macrophage (green arrowhead). Sections were stained for ACE2 using DAB and counterstained with hematoxylin.
We identified 12 lung samples from 11 patients requiring mechanical ventilation for AHRF available from our institution’s archive. All 11 patients had DAD (Suppl. Table 2), a pathological finding consistently observed in patients with severe Covid-19 lower respiratory disease10,11. All tissue was collected prior to 2019 (range 2010 – 2018), excluding the possibility of SARS-CoV-2 involvement. H&E control sections revealed the expected histologic findings, depending on the phase of DAD, with reactive AT2 hyperplasia, hyaline membrane formation, interstitial thickening and/or fibrosis, and exudative edema (Suppl. Figure 1B). When stained by IHC, prominent ACE2 expression within the alveolar parenchyma was observed in reactive AT2 cells with increasing ACE2 staining intensity detected with advanced age (Figure 3A and 3B). ACE2 expression was quantified either by normalizing to tissue area (Figure 3C) or cellularity (Figure 3D) which revealed a significant increase with age (p = 0.004 and p = 0.003, respectively, linear regression). As a control for precision, 1 of the 11 patients had a double lung explant with 2 samples collected contemporaneously (1 from the left lung and 1 from the right lung). In this 67 year-old-man, the pathological findings were the same for each sample (Suppl. Table2). Importantly, both specimens yielded similar results when quantified for ACE2, indicating that our procedure measured its expression with good intra-individual reproducibility (Figure 3C and 3D). Excluding either of these samples did not change the effect of age on ACE2 expression (p = 0.009 by area and p = 0.003 by cellularity excluding the right lung; p = 0.008 by area and p = 0.006 by cellularity excluding the left lung, linear regression).

A) Representative images of lung stained for ACE2 from a 67-year-old man with acute lung injury superimposed on fibrosing interstitial lung disease are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). Numerous reactive AT2 cells (red arrowheads) exhibiting nucleomegaly and abundant cytoplasm can be seen demonstrating strong ACE2 staining. B) Representative images of lung stained for ACE2 from a 40-year-old man with acute lung injury are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). Clusters of reactive AT2 cells (red arrowheads) are present along the alveolar septum which exhibit low level ACE2 expression. C) Quantitative IHC for ACE2 was carried out on samples from ventilated patients (n = 12 samples from 11 patients). Total ACE2 expression from 5 low power fields is plotted relative to the patient’s age at the time of specimen collection. A linear fit to the data is indicated by the dashed line with the 95% confidence interval highlighted in grey. D) The same specimens quantitated in (C) were normalized by cellularity and the average ACE2 expression per cell is plotted along with a linear fit to the data and its 95% confidence interval. The red arrow in (C) and (D) indicates a patient providing 1 sample from the left lung and 1 sample from the right lung during the same procedure, utilized as a control for intra-individual reproducibility. Sections were stained for ACE2 using DAB and counterstained with hematoxylin.
Alveolar macrophages comprised a minor fraction of total cells in samples from ventilated patients (median 3.4%, range 1.7% - 10.9%) and by visual inspection did not show the prominent age-related change in ACE2 expression seen in AT2 cells. To determine whether the abundance of alveolar macrophages contributes strongly to the effect of age on ACE2 expression, we quantified the number of alveolar macrophages in each sample. When normalized to either tissue area (Suppl. Figure 5A) or cellularity (Suppl. Figure 5B), the number of alveolar macrophages did not change with age (p = 0.768 and p = 0.427, respectively, linear regression).
To compare our results in ventilated patients to samples collected from spontaneous breathing individuals, we collected an additional set of 20 lung samples from patients who underwent lung excision/biopsy for either pneumothorax repair or metastatic non-pulmonary neoplasms (Suppl. Table2). In all cases, care was taken to select portions of adjacent uninvolved lung parenchyma to analyze for ACE2 expression by IHC. Similar to our control sections, ACE2 staining was largely found within AT2 cells and alveolar macrophages (Figure 4A and 4B). When compared across ages, there was no significant difference either upon visual inspection or when quantified normalizing to tissue area (p = 0.231, linear regression) (Figure 4C) or cellularity (p = 0.349, linear regression) (Figure 4D).
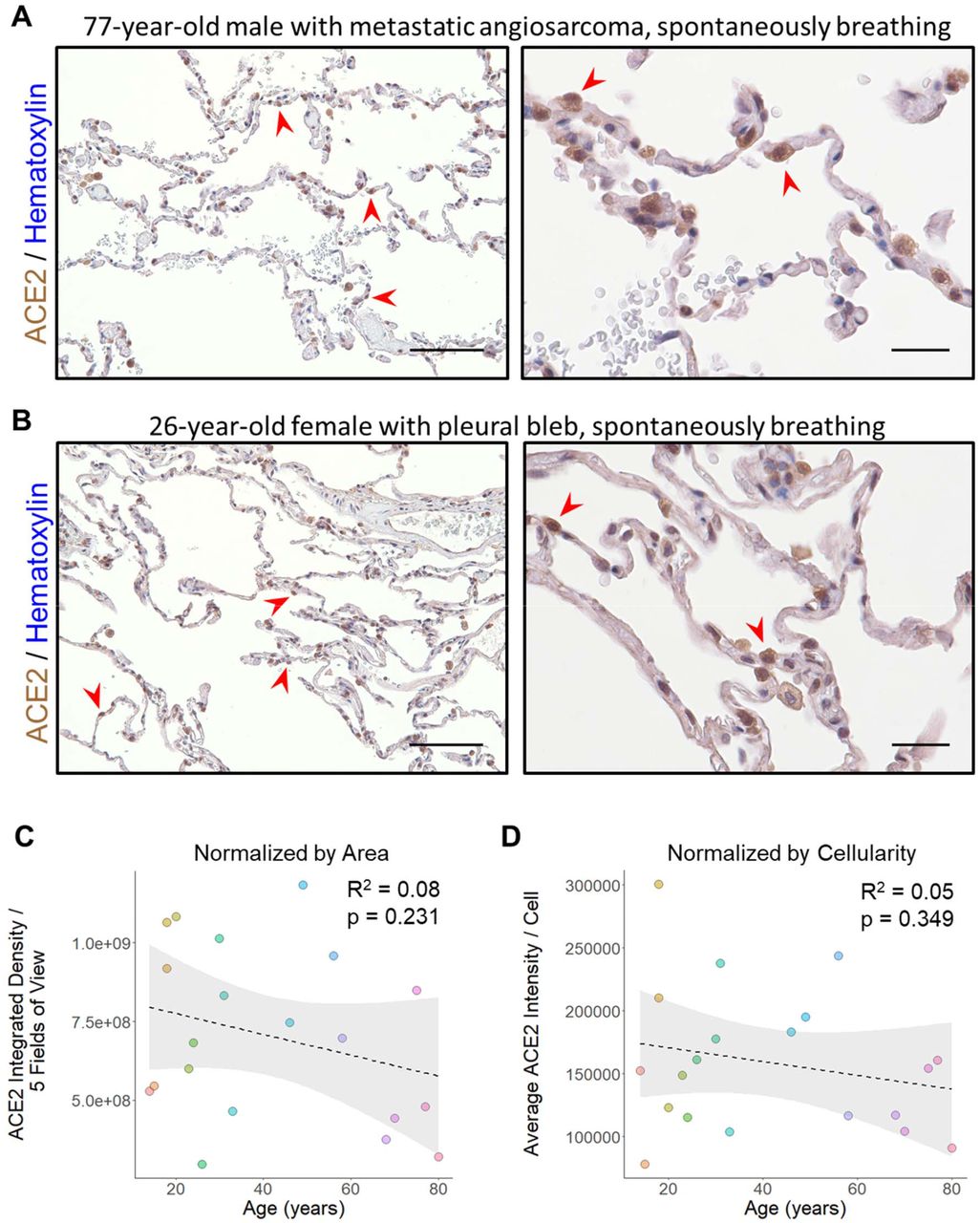
A) Representative images of lung stained for ACE2 from a 77-year-old man undergoing wedge resection for suspected metastatic angiosarcoma are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). The normal alveolated lung parenchyma adjacent to the lesion reveals AT2 cells which are positive for ACE2 (red arrowheads) scattered among largely ACE2 low type I pneumocytes. B) Representative images of lung stained for ACE2 from a 26-year-old female who underwent blebectomy for a spontaneous pneumothorax are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). Histologic findings were similar to those found in (A) with strong ACE2 staining in scattered AT2 cells (red arrowheads) and occasional alveolar macrophages. C) Quantitative IHC for ACE2 was carried out on samples from non-ventilated patients (n = 20 patients). Total ACE2 expression from 5 low power fields is plotted relative to the patient’s age at the time of specimen collection. A linear fit to the data is indicated by the dashed line with the 95% confidence interval highlighted in grey. D) The same specimens quantitated in (C) were normalized by cellularity and the average ACE2 expression per cell is plotted along with a linear fit to the data and its 95% confidence interval. Sections were stained for ACE2 using DAB and counterstained with hematoxylin.
During our image acquisition, we noticed 2 cases exhibiting prominent ACE2 expression within the lung vascular endothelium (Figure 5A and 5B). This feature was not observed in any of the other 29 cases we examined. Upon chart review, we discovered that the first case was from an 83-year-old man receiving valsartan during his immediately preceding admission at an outside institution, and at the time of transfer to our hospital, where his surgical resection occurred on post transfer day 4 (Figure 5A). The second case was from a 53-year-old man who was actively receiving inpatient lisinopril at the time of lung explant (Figure 5B). Both patients were on supportive mechanical ventilation for AHRF prior to lung sample collection. The prominent age-related changes in AT2 cell ACE2 expression were again evident, but in addition, there was strong staining within the endothelium, indicating increased ACE2 expression within the vasculature. None of the other 29 cases in our dataset were from patients on angiotensin-converting enzyme inhibitors (ACEIs) or angiotensin receptor blockers (ARBs) (Suppl. Table2).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A) Representative images of lung stained for ACE2 from an 83-year-old man in the organizing phase of diffuse alveolar damage are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). In addition to high ACE2 expression in reactive AT2 cells (green arrowheads), strong staining can be seen in numerous vascular endothelial cells (red arrowheads) throughout the thickened septal area in this patient on valsartan 4 days prior to specimen collection. B) Representative images of lung stained for ACE2 from a 53-year-old man with organizing diffuse alveolar damage superimposed on fibrosing interstitial lung disease are shown at low power (left, scale bar 200µm) and high power (right, scale bar 50µm). AT2 cell hyperplasia can be seen with lower level ACE2 expression present along the alveolar septum (green arrowheads). Strongly stained endothelial cells (red arrowheads) can be seen throughout the entire specimen from this patient who was on daily lisinopril at the time of specimen collection. Sections were stained for ACE2 using DAB and counterstained with hematoxylin.
Discussion
After the recognition that SARS-CoV-2 depends on ACE2 for host infection23, a number of key studies have linked the expression of this viral receptor to tissue susceptibility. Within the mammalian body, expression of ACE2, along with transmembrane serineprotease 2 (TMPRSS2)--the human protease responsible for activating the SARS-CoV-2 spike protein--coincides with cell types involved by Covid-1939. ACE2 protein is highly expressed in differentiated gut enterocytes, leading to productive infection with SARS-CoV-240. Interestingly, ACE2 expression is upregulated in the airway and lungs of smokers29, a potential risk factor for severe disease41. Additionally, children have lower ACE2 RNA expression in their nasal epithelium compared with adults42. A growing body of evidence suggests that ACE2 is highly associated with disease susceptibility and severity. Given that the mechanism of death in Covid-19 typically involves severe lower respiratory tract infection, a disease feature strongly correlated with age, previous studies have sought to connect lung ACE2 expression with aging28,29. Although we analyzed a subset of the same publicly available data, we highlight the critical impact of mechanical ventilation on ACE2 expression, an important physiologic change for the lung. By incorporating this feature into our model, we find that ACE2 expression indeed increases in older individuals through what is likely a combination of advanced age, and either the need for or consequence of mechanical ventilation.
Reactive AT2 hyperplasia has long been recognized as a common finding after alveolar epithelial damage43–46. Patients in our ventilator supported cohort uniformly exhibited DAD with abundant hypertrophic AT2 cells. Our IHC analyses reveal that AT2 cells, as well as alveolar macrophages, are the primary cell types expressing ACE2 in the alveolus. Although we cannot rule out a subtle shift, changes in ACE2 expression among alveolar macrophages was minor when compared visually to AT2 cells and the former’s abundance showed no relationship with age in ventilated patients. Rather, the age-related increase in ACE2 expression we observed appears to be primarily mediated by the level of this protein’s expression in individual AT2 cells, with a potential lower contribution from other cells types. This trend was not seen in spontaneous breathing patients suggesting that the response of AT2 cells to alveolar injury drives this change. ACE2 expression in AT2 cells has been shown to dramatically decrease upon cell cycle entry47. Furthermore, a recent study has revealed that ACE2 expression is upregulated in human respiratory cells in response to inflammatory signals including IFNα2 and IFNγ39. Immune changes associated with aging include the chronic, low-grade endogenous production of inflammatory cytokines, frequently referred to as “inflammaging”, which is caused by a number of processes48–51. One potential model for the age-related differences we observe would be that in response to alveolar damage, AT2 cells enter the cell cycle and ordinarily downregulate ACE2 expression. However, in older individuals, the increase in circulating inflammatory cytokines prevents this downregulation or even leads to ACE2 upregulation alongside AT2 cell proliferation. Alternatively, epigenetic dysregulation may preclude the proper downregulation of ACE2 in aged AT2 cells22. This model implies that the relationship of aging to ACE2 expression is associated with the need for mechanical ventilation (i.e., the alveolar damage itself) instead of being caused by it. However, the reverse scenario also merits consideration.
Positive pressure ventilation is associated with lung injury (ventilator-associated lung injury or VALI) and is sometimes the underlying cause of such injury (ventilator-induced lung injury or VILI)52. The pathological features of VILI are indistinguishable from other causes of ARDS and are induced by volume and pressure stress within the alveolus53. Ventilation strategies that result in VILI provoke higher levels of inflammatory cytokine release54,55. In animal models, advanced age is associated with an increased likelihood of VILI56 and exacerbated cytokine release in response to ventilation57. If lung ACE2 expression is triggered by mechanical ventilation itself via inflammatory cytokine production, then these factors may predispose older individuals to severe SARS-CoV-2 infection. The relative contributions of these and other potential mechanisms remain to be explored.
Although our study was originally designed to characterize the effects of age and ventilation on ACE2 expression, during data acquisition, the presence of strong vascular ACE2 staining in 2 cases was striking. This prompted us to investigate the underlying cause. Chart review uncovered that 1 patient was receiving an ARB just prior to sample collection and the second was receiving an ACEI at the time of collection. The remaining 29 cases were taken from patients not receiving either class of medication. ACE2 has been shown previously to be upregulated by ACEI/ARB therapy in the rodent heart58 and human intestine59, raising the concern that these therapeutics may predispose to SARS-CoV-2 infection60. Observational studies have indicated that the use of ACEI/ARBs does not increase the chances of testing positive for SARS-CoV-261 and reduces the chances of Covid-19 mortality62. Randomized trials to assess the impact of ACEI/ARB discontinuation in Covid-19 are currently planned and ongoing63,64. Although we identified elevated pulmonary endothelial ACE2 expression in the only 2 patients on ACEI/ARBs in our cohort, this finding is preliminary and requires replication. A recent study has demonstrated pulmonary vascular endothelialitis associated with SARS-CoV-2 infection of these cells9. Whether increased ACE2 expression modulates this disease feature or Covid-19 pathogenesis in general warrants further investigation.
Limitations
In this small series of cases, we identified increased ACE2 RNA and ACE2 protein expression within the human lung associated with age, when controlling for ventilator status. Although our RNAseq analyses drew on the sample rich resource of the GTEx dataset, including 578 unique donor lung samples, our quantitative IHC results derived from limited archival samples (12 samples from 11 patients). However, the concordance between our findings at the RNA and protein level revealing an interaction of mechanical ventilation and age in both cases, strengthens our conclusions.
Data Availability
The manuscript utilizes publicly available data from the GTEx Portal and the representative data presented in the submitted files.
Author Contributions
S.A. Baker, G.J. Berry, and T.J. Montine conceived the study and wrote the manuscript. S.A. Baker and S. Kwok obtained the data and performed the analysis. S.A. Baker, S. Kwok, G.J. Berry, and T.J. Montine have no conflicts of interest to declare
Acknowledgments
We would like to thank the staff of the Stanford Health Care Department of Pathology for excellence in patient care throughout the current pandemic and selfless diligence in cataloguing, processing, and reporting the original case material contributing to this study.
References



