
Abstract
Background Early in the COVID-19 pandemic type 2 diabetes (T2D) was marked as a risk factor of severe disease and mortality. Inflammation is central to the aetiology of both conditions where variations in immune responses have the potential to mitigate or aggravate disease course. Identifying at risk groups based on immuno-inflammatory signatures is valuable in directing personalised care and developing potential targets for precision therapy.
Methods This study characterised immunophenotypic variation associated with COVID-19 severity in type 2 diabetes. Broad-spectrum immunophenotyping quantified 15 leukocyte populations in peripheral circulation from a cohort of 45 hospitalised COVID-19 patients with and without type 2 diabetes.
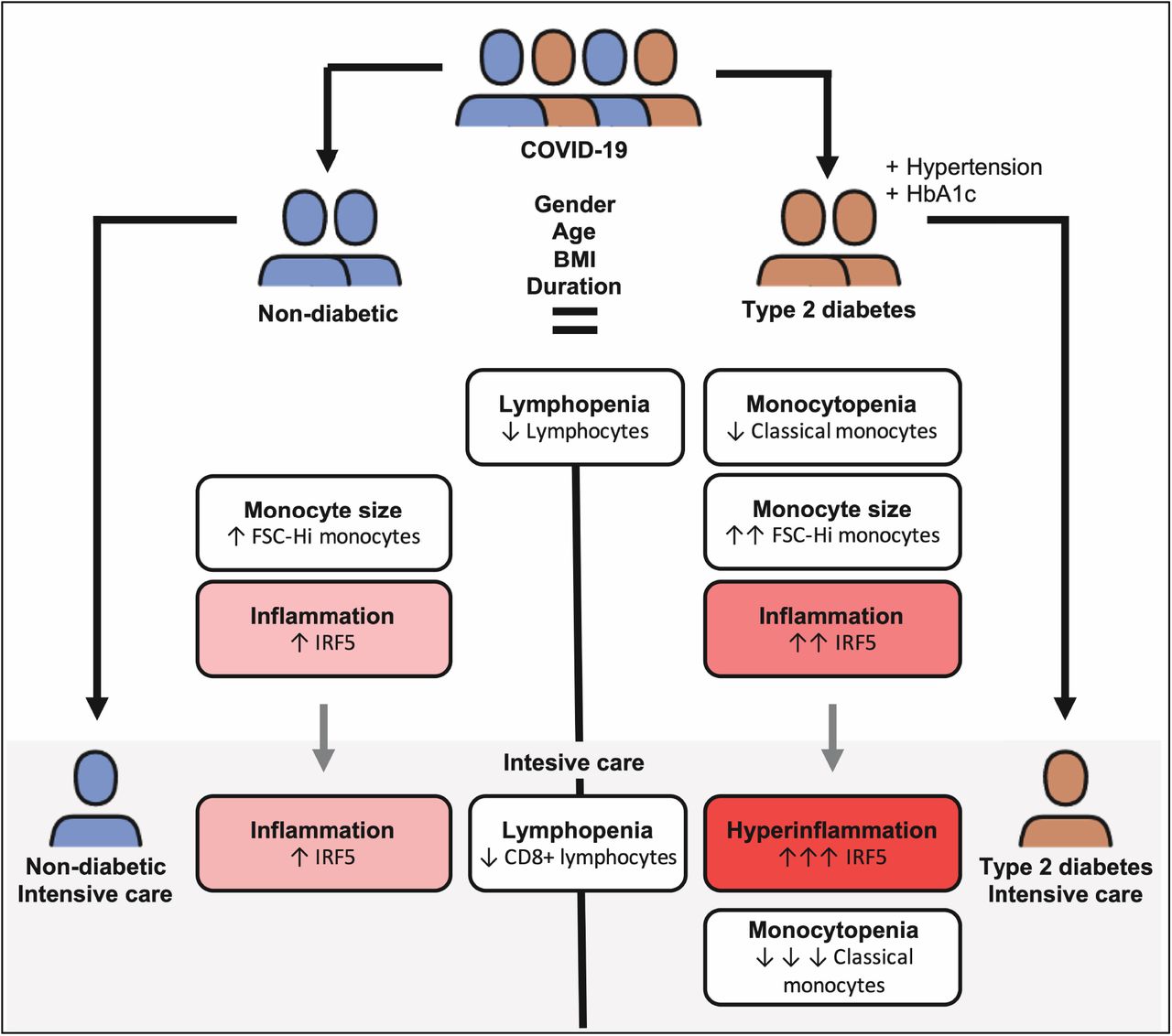
Results Morphological anomalies in the monocyte pool, monocytopenia specific to quiescent monocytes, and a decreased frequency of cytotoxic lymphocytes were associated with severe COVID-19 in patients with type 2 diabetes requiring intensive care. An aggravated inflammatory gene expression profile, reminiscent of the type-1 interferon pathway, underlaid the immunophenotype associated with severe disease in T2D.
Conclusion Shifts in T-cell and monocyte dynamics underpin a maladaptive response to SARSCoV-2. These alterations may impact type-1 interferon signalling which is the likely source of the hyperinflammation that increases voracity of COVID-19. These findings allow the identification of type 2 diabetic patients at risk of severe disease as well as providing evidence that the type-1 interferon pathway may be an actionable therapeutic target for future studies.
Trial registration NCT02671864
Funding French National Agency of Research (ANR); European Foundation for the study of diabetes (EFSD); European Research Council (ERC); Francophone Society for Diabetes (SFD)
Brief summary Maladapted monocyte responses including class switch, morphological anomalies and systemic hyperinflammation put patients with type 2 diabetes at higher risk of severe COVID-19
{kind=link}
Competing Interest Statement
The authors have declared no competing interest.
Clinical Trial
NCT02671864
Funding Statement
Funding: French National Agency of Research (ANR); European Foundation for the study of diabetes (EFSD); European Research Council (ERC); Francophone Society for Diabetes (SFD)
Author Declarations
I confirm all relevant ethical guidelines have been followed, and any necessary IRB and/or ethics committee approvals have been obtained.
Yes
The details of the IRB/oversight body that provided approval or exemption for the research described are given below:
This study was approved by local institutions and ethical committees, the Ethics Committee of CPP Ile-de-France granted approval for all individuals (Ile de France V number 15070). All patients provided informed consent indicating that they understood the nature of their participation in the study (NCT02671864). The principal investigator of this clinical trial is Prof. Gautier Jean-Francois: jean-francois.gautier@aphp.fr.
All necessary patient/participant consent has been obtained and the appropriate institutional forms have been archived.
Yes
I understand that all clinical trials and any other prospective interventional studies must be registered with an ICMJE-approved registry, such as ClinicalTrials.gov. I confirm that any such study reported in the manuscript has been registered and the trial registration ID is provided (note: if posting a prospective study registered retrospectively, please provide a statement in the trial ID field explaining why the study was not registered in advance).
Yes
I have followed all appropriate research reporting guidelines and uploaded the relevant EQUATOR Network research reporting checklist(s) and other pertinent material as supplementary files, if applicable.
Yes
Data Availability
Information with regards to data availability on request. Prior to peer-review raw data cannot be made public. We conform to standard requirements.



